
Alan Wadei
Honey bees of the Apis genus are eusocial insects, that is they are socially highly-organised raising brood cooperatively and intergenerationally, and are specialised in their division of labour. They range naturally across Africa, Europe and Asia but are absent at high latitudes. This natural distribution excludes the Americas and the Pacific nations of the New World: the original distribution of Apis mellifera and Apis cerana is shown in Figure 1ii.
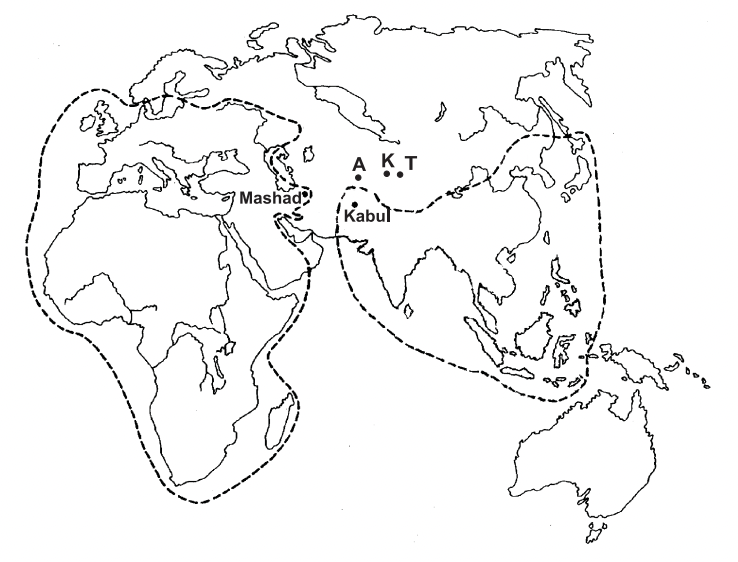
Figure 1 Native range of the Western Honey Bee (Apis mellifera) [left Africa-Europe] and of the Asian Honey Bee (Apis cerana)[right Asia-China-USSR]
Biogeography of Apis species
The honey bees of the Apis genus are wide-spread. Apis mellifera, Apis cerana and Apis dorsata have a number of geographically delineated races and halotypes, their differences are supported by both morphometric and genetic studies. The most geographically restricted species Apis koschevnikovi, Apis nigrocinta and Apis nuluensis (or Apis cerana nuluensis) have diverged from Apis cerana as separate species in relatively recent times. Much also remains to be understood about the native range of some of the Asian and African honey bee subspecies. For example, the distribution of Apis nigrocincta (the Philippine Honey Bee) has been inferred from just a few surveys.
The greatest biodiversity of honey bee species is in the South China Sea region in and around the Phillipines, Kalimantan and the Sulawesi and is where the ancestral genus appears likely to have radiated from.
With the rise of agrarian societies during the Holocene (the geological epoch spanning the last 11,500 years), honey bees have been transported far and wide: both Apis mellifera and Apis cerana have been employed extensively for pollination and honey production. The contemporary range of the Apis genus is shown in Figure 2. While many social bee species, including Apis species (other than A. cerana and A. mellifera), have also been exploited for both honey production and pollination services, their geographic range is typically contracting due to habitat destruction and modification. Some Apis species, as well as Apis subspecies, have a very high swarming and absconding propensity. Together, these factors have limited their usefulness and, in some cases, threatened their survival.
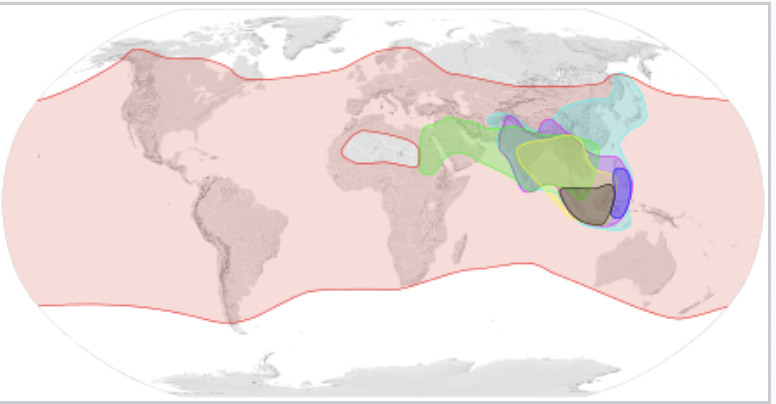
Figure 2 Apis distribution map showing contemporary distribution of dwarf, medium-sized and giant honey bees:iii
Apis andreniformis–—
Apis florea—-
Apis cerana—-
Apis koschevnikovi—-
Apis nigrocincta—-Apis mellifera—-
Apis dorsata—-
Natural range of species and subspecies of honey bees
Honey bees of the modern era fall into three subgenera, the less socially complex giant honey bees (Megapis) that nest on a single exposed comb, the advanced cavity dwelling honey bees (Apis) that include Asian and Western honey bees on multiple combs and the least socially evolved dwarf honey bees (Micapis) also restricted to a single comb. Both Apis cerana and Apis mellifera extended their natural range into temperate and arid climes by taking advantage of their sheltered nesting habit, by hoarding and storing honey (carbohydrate) and pollen (protein) in brief times of abundance and by complex absconding and reproductive swarming behaviours.
The contemporary delineation of honey bee species and subspecies is based on morphometric and genetic studies that take into account their geographic origins. Cervanciaiv describes eight species of honey bees: Apis florea Fabricius, Apis andreniformis Smith, Apis cerana Linnaeus, Apis nuluensis Tingek, Koeniger and Koeniger, Apis nigrocincta Smith, Apis mellifera Linnaeus, Apis dorsata Fabricius and Apis laboriosa Smith. Apis koschevnikovi Buttel-Reepen must be added to this listing as must also now be added the Indian Plains Honey Bee (Apis indica) and the giant Phillipine Honey Bee (Apis breviligula) though a further giant bee honey species from the Sulawesi (currently designated Apis dorsata binghami) remains in doubt. There is some evidence that Apis nuluensis Tingek, Koeniger and Koeniger is a separate taxon and may have recently diverged from A. ceranav.
So how many distinct honey bee species are there? Arias and Sheppardvi suggest there is sufficient phylogenetic evidence to indicate that there are ten distinct living species, giant bees (Apis dorsata, Apis binghami and Apis laboriosa), dwarf bees (Apis andreniformis and Apis florea), and cavity-nesting bees (Apis mellifera, Apis cerana, Apis koschevnikovi, Apis nuluensis and Apis nigrocincta).
With the more recent addition of the Giant Philippine Honey Bee (Apis breviligula) and the Indian Plains Honey Bee (Apis indica)vii we might conclude that there are as many as a dozen types of honey bee, the main semantic dispute lying with the number of Apis cerana related species that have split off and the uncertain status of the Giant Sulawesi Honey Bee (Apis dorsata binghami) still considered by most to be a sub species of Apis dorsata. As I’m neither a taxonomist, a molecular geneticist nor an evolutionary biologist, I’ve simply come up with a candidate listing (see Table 1 in my more recent review of Giant Honey Bees: next essay) where I conclude that there seem to be 10-12 separate taxa.
Honey bees harbour a number of parasitic arachnids reflecting their geographic origins: some have spread to honey bee subspecies while others have jumped species: Ascarapis woodi, Varroa destructor, Tropilaelaps clareae and Tropilaelaps mercedesae have done so with devastating consequences:
Euvarroa has evolved with the dwarf honey bees, Apis andreniformis and Apis florea;
Varroa has evolved with several subspecies of Apis cerana;
Tropilaelaps is associated with different subspecies of the giant honey bee Apis dorsata; and
Ascarapis origins are not clearviii. Acarapis species affect honey bees worldwide but their origins and co-evolution with bees do not appear to have been elucidated: Acarapis woodi (Acarine Mite) is known from its association with Apis mellifera and is now also a parasite of Apis cerana.
Bacterial, fungal, microsporidian, protozoan and viral diseases affecting Apis species are of obscure origin and their association with hymenoptera (bees, wasps and ants) are the subject of active investigation and are not reviewed here.
The dwarf honey bees (subgenus Micapis)
There are two dwarf honey bees, the more western and the more widely distributed Apis florea and the more eastern Apis andreniformis: Apis andreniformis tends to live at higher altitudes where the species are sympatric, that is where their ranges overlap.
Apis florea (Host to the parasite Euvarroa sinhai, that has also been reported on A. mellifera imported to an area where Apis florea is present but does not reproduce on the Western Honey Bee) – continental Asia and Africa, southeastern Asia: Thailand, Iran, Oman, India, Myanmar, and some parts of China, Cambodia, and Vietnam and forested regions of the Middle East (Figure 3).
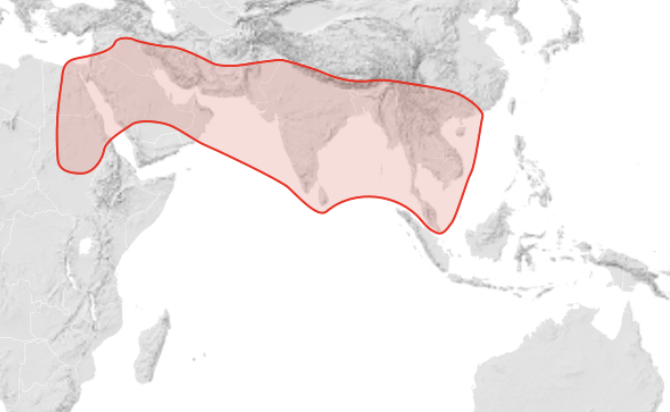
Figure 3 Apis florea distributionix
Apis andreniformis (This honey bee is parasitised by both Euvarroa wongsirii and Euvarroa sinhai) – Southeast Asia: southern China, India, Burma, Laos, Vietnam, Thailand, Malaysia, Indonesia, and the Philippines (Figure 4).
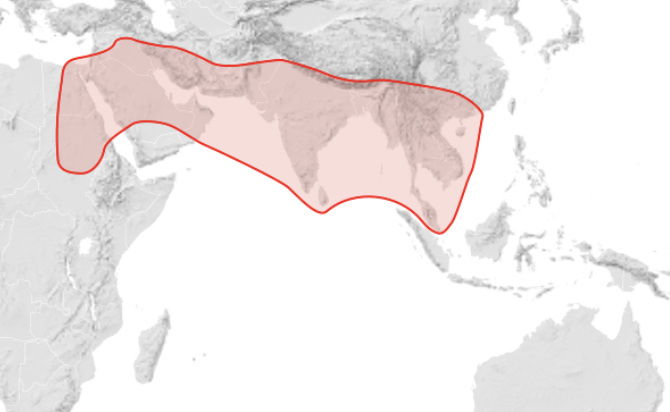
Figure 4 Apis andreniformis distributionx
Medium-sized, cavity-dwelling honey bees (subgenus Apis)
The medium-sized, cavity-dwelling species comprise a cluster of four or five species. The best known and widely distributed Apis cerana and Apis mellifera are of common, if disputed evolutionary origin. Many of their nest structural, pheromonal and behavioural characteristics are shared, although there are many subtle differencesxi. Two Three, perhaps four geographically distinct restricted species, Apis indica, Apis koschevnikovi and Apis nigrocincta, (also Apis nuluensis, if it is regarded as a separate taxon), are very closely allied to Apis cerana. This species complex has been well researched and the breakdown of subspecies follows essentially that of Engelxii.
Apis cerana Asian Honey Bee(Parasites include the microsporidian Nosema ceranae; and the arachnids Varroa destructor (two genotypes have crossed over to Apis mellifera), Varroa jacobsonixiii and Ascarapis woodi – the Western Honey Bee Tracheal Mite) – is widely distributed with eight, geographic subspecies (Figure 5):
Apis cerana cerana
Apis cerana heimifeng
Apis cerana indica
Apis cerana japonica
Apis cerana javana
Apis cerana johni
Apis cerana nuluensis
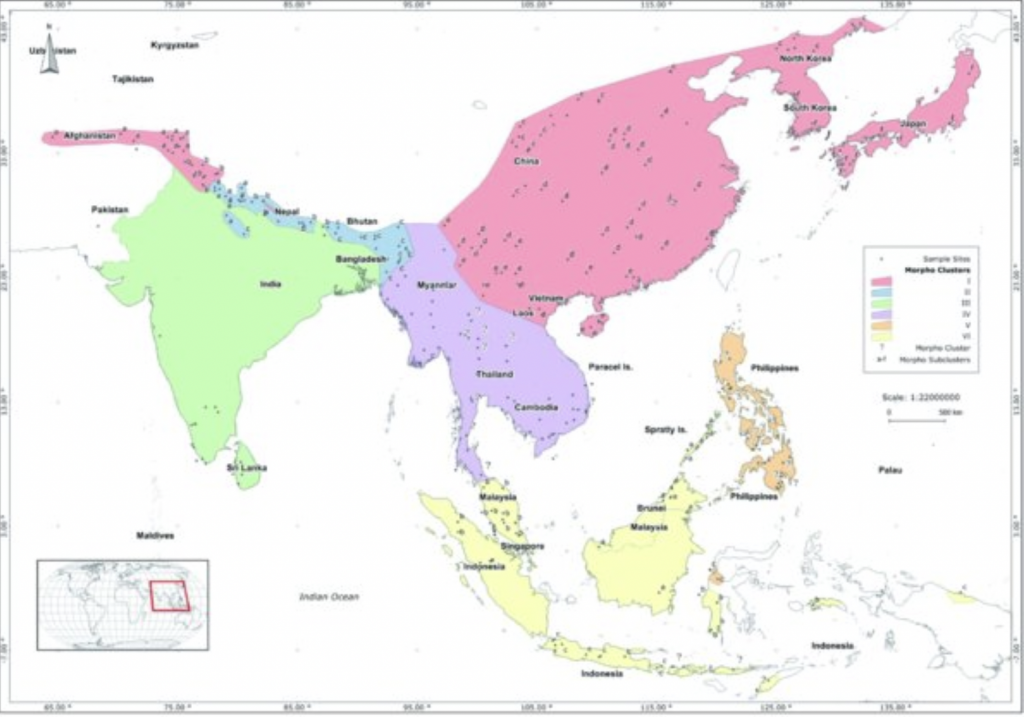
Figure 5 Morphometric clusters of the Asian Honey Bee, Apis ceranaxiv
Their distribution, like that of Apis mellifera, is remarkable:
Apis cerana cerana (the Asian Honey Bee) – Afghanistan, Pakistan, north India, and south along the central deserts and mountain ranges, across most of central and southern China, along the eastern edge of Asia up to Korea and Ussuria and south to Northern Vietnam.
Apis cerana heimifeng (the Black Chinese Honey Bee) – Central China at relatively high elevation: northern Sichuan Province, southwestern Gansu Province, and Eastern Qinghai Province.
Apis cerana indica – (the Indian Honey Bee) South India, Sri Lanka, Bangladesh, Burma, Malaysia, Indonesia and the Philippines.
Apis cerana japonica – (the Japanese Honey Bee) – Japan, Thailand.
Apis cerana javana (the Javanese Honey Bee) – Java to Timor, now also PNG, Irian Jaya and NE Australia.
Apis cerana johni (the Sumatran Honey Bee) – Sumatra.
Apis cerana nuluensis (the Malaysian Mountain Honey Bee) – Mountains of Sabah in east Malaysia and Kalimantan (Figure 6). (This particular bee, together with Apis nigrocincta, is host to Varroa underwoodi).
Apis cerana skorikovi (the Himalayan Honey Bee) – Central and east Himalayan mountains (Himalayan Uplift).
Apis indica (Plains Honey Bee of southern India)
Long regarded as a subspecies of Apis cerana, the Plains Honey Bee of southern India has only recently been accorded separate species status, largeley based on its genetic makeupxv. Formerly classified as Apis cerana indica, its range extended from South India, Sri Lanka, Bangladesh, Burma, Malaysia, Indonesia (presumably excluding Java and the archipelago east) and the Philippines.
Apis nuluensis (Malaysian Mountain Honey Bee)
The Malaysian Mountain Honey Bee is restricted to a mountainous region of Sabah (1400-4100 m) in the state of Sabah in East Malaysia (Figure 6) . (This particular bee, together with Apis nigrocincta, is host to Varroa underwoodi).
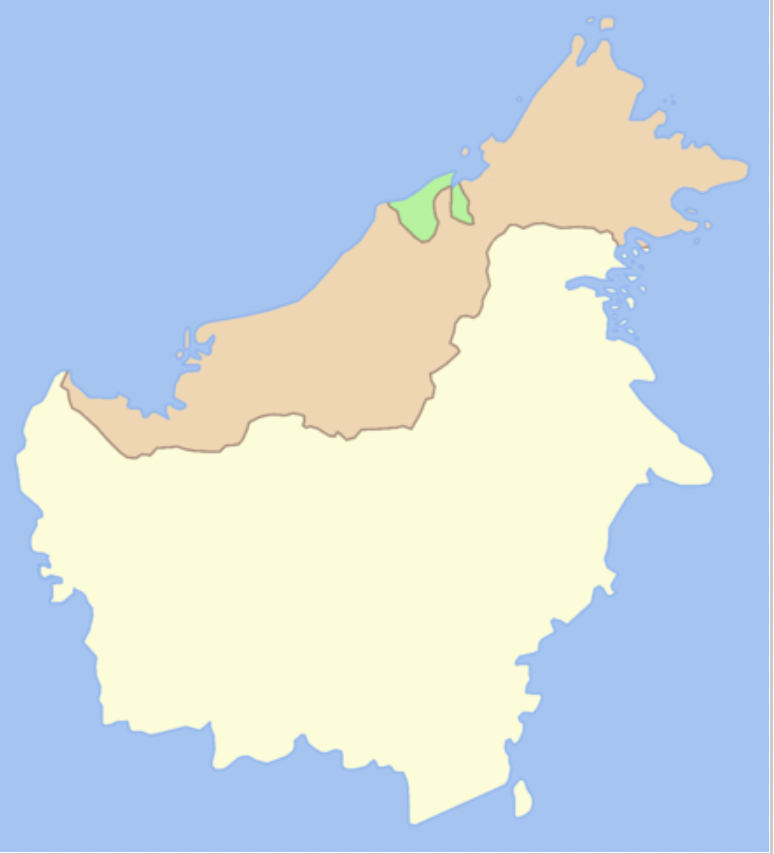
Figure 6 Apis cerana nuluensis distributionxvi
Apis nigrocinta (the Philippine Honey Bee) – Sulawesi and the nearby islands of Sangihe, and the Phillipines (Mindanao) (Figure 7). (Apis nigrocinta is parasitised by Varroa underwoodi.)
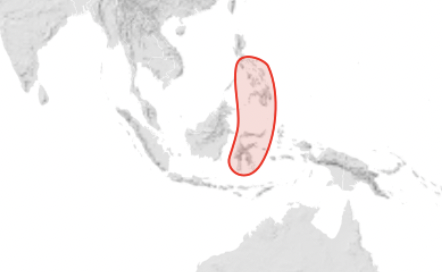
Figure 7 Apis nigrocincta distributionxvii
Apis koschevnikovi (Koschevnikovi’s Honey Bee) (Figure 8). (This species is the natural host of Varroa rindereri. However this parasite has spread to Apis cerana and is now in Irian Jaya and PNGxviii) – Malaysian Peninsular, Sumatra and also Kalimantan where it is sympatric with Apis cerana nuluensis (Apis nuluensis).
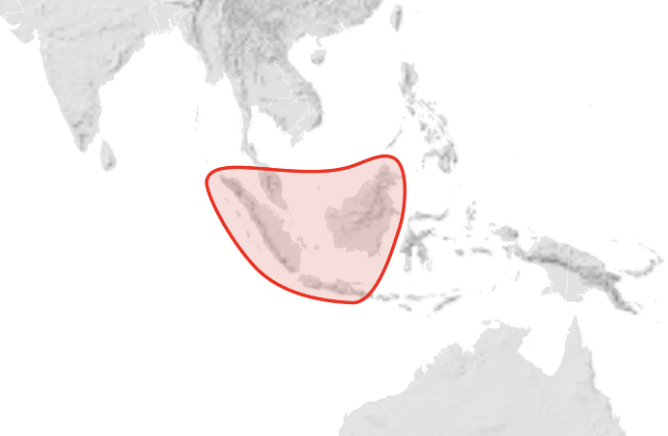
Figure 8 Apis koschevnikovi distributionxix
Apis mellifera (Western Honey Bee) (Natural parasites include the microsporidian Nosema apis and the arachnid Ascarapis woodi). – The Western Honey Bee is widespread in the Old World being found naturally in Africa, the Middle East and Europe.
There are twenty eight, mainly geographically isolated, subspecies:
Apis mellifera adami
Apis mellifera adansonii
Apis mellifera anatoliaca
Apis mellifera artemsia
Apis mellifera capensis
Apis mellifera carnica
Apis mellifera caucasia
Apis mellifera cecropia
Apis mellifera cypria
Apis mellifera iberiensis
Apis mellifera intermissa
Apis mellifera jemenitica
Apis mellifera lamarckii
Apis mellifera ligustica
Apis mellifera litorea
Apis mellifera macedonica
Apis mellifera meda
Apis mellifera mellifera
Apis mellifera monticola
Apis mellifera pomonella
Apis mellifera remipes
Apis mellifera ruttneri
Apis mellifera sahariensis
Apis mellifera scutellata
Apis mellifera siciliana
Apis mellifera sosimii
Apis mellifera syriaca
Apis mellifera unicolor
Whitfield and co-workers have described the origins of the Western Honey Bee (Apis mellifera) and its European, Asian and African races classifying them into four branchesxx based on the pioneering morphological studies of Ruttner and co-workers (Figure 9)xxi. A follow up study by Cridland, Tsutsui and Ramírexxii further elucidates the origins of the Western Honey Bee and builds on this hypothesis.
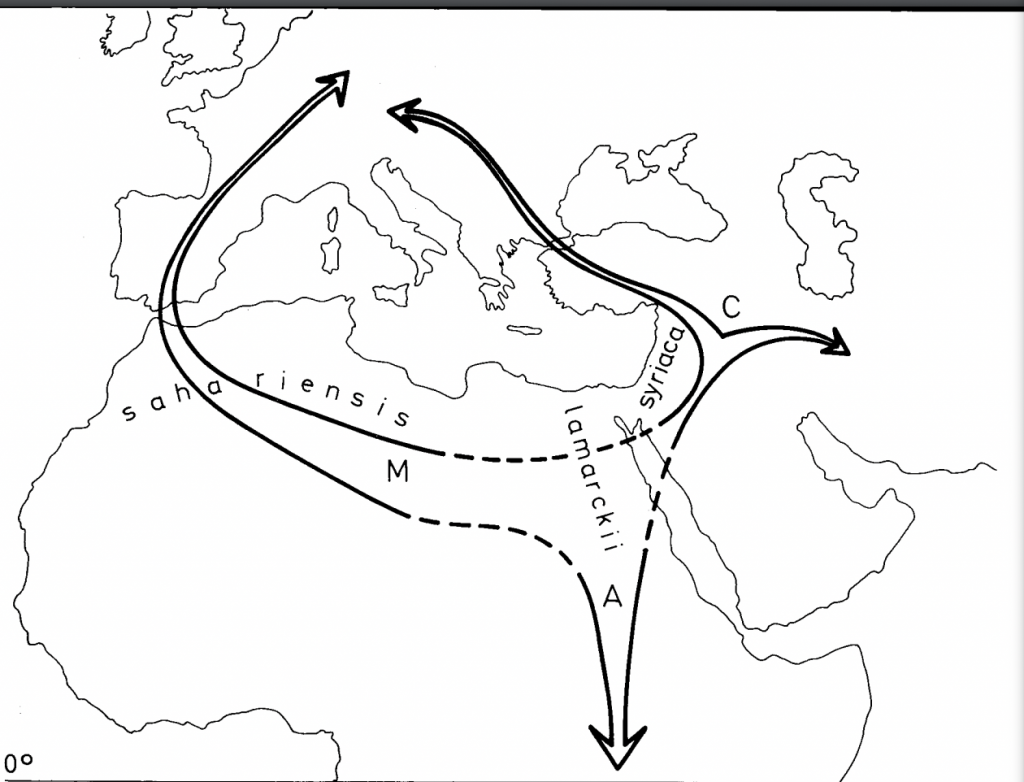
O
Figure 9 Morphometric clusters of the Western Honey Bee subspecies, Apis mellifera
African subspecies (branch A), northwestern European subspecies (branch M), southwestern European subspecies (branch C) and Middle-Eastern subspecies (branch O).
Northern European subspecies
Apis mellifera mellifera(the German Black Bee or the European Honey Bee) – central Europe north of the Alps to western southern France, southern Sweden, British Isles
Southern European subspecies
Apis mellifera artemsia(the Russian Steppe Honey Bee) – Central Russian Steppes
Apis mellifera carnica (the Carniolan Honey Bee) – Carniola region of Slovenia, the Eastern Alps and northern Balkans: Yugoslavia, Romania
Apis mellifera cecropia (the Greek Honey Bee) – Greece and surrounding Aegean Islands
Apis mellifera iberiensis (the Iberian Honey Bee) – Iberian Península (Spain and Portugal)
Apis mellifera ligustica (the Italian Honey Bee) – Italian Peninsular
Apis mellifera macedonica (the Macedonian Honey Bee) – Northern Greece (Macedonia and Thrace), Republic of Macedonia
Apis mellifera ruttneri (the Maltese Honey Bee) – Maltese Islands
Apis mellifera siciliana (the Sicilian Honey Bee) – Sicily
Apis mellifera sosimii (the Ukrainian Honey Bee) – Ukraine and northern Caucasus Mountains
Middle Eastern and Asian subspecies
Apis mellifera adami (the Cretian Honey Bee) – Crete
Apis mellifera anatoliaca (the Anatolian Honey Bee) – central region of Anatolia in Turkey and Iraq extending as far east as Armenia
Apis mellifera caucasia (the [grey] Caucasian Honey Bee) – the Caucasus Mountains
Apis mellifera cypria (the Cyprian Honey Bee) – Cyprus
Apis mellifera meda (the Median Honey Bee) – Iran, Iraq, southern Turkey and Northern Syria
Apis mellifera pomonella (the Tien Shan Honey Bee) – Tien Shan Mountains in Central Asia
Apis mellifera remipes (the Yellow Armenian Honey Bee) – Armenia (This subspecies is similar to and possibly the same as Apis mellifera intermissa.)
Apis mellifera syriaca – (Syrian Honey Bee) Near East and Israel
African subspecies
Apis mellifera adansonii (the West African Honey Bee) – Niger, Senegal, Zaire
Apis mellifera capensis – (the Cape Honey Bee) South Africa
Apis mellifera intermissa (the Tellian Honey Bee) – northern coast of Africa as far west as Morocco, Libya, Tunisia bordered by the Atlas Range
Apis mellifera jemenitica (the Arabian or Nubian Honey Bee) – Somalia, Uganda, Sudan, Yemen
Apis mellifera lamarckii – (Lamarck’s Honey Bee) – Nile valley: Egypt and Sudan
Apis mellifera litorea – Low elevations of east Africa – e.g. Kenya
Apis mellifera monticola(the East African Mountain Honey Bee) – High altitude mountains of East Africa:Kenya, Tanzania: Mt Elgon, Mt Kilimanjaro, Mt Kenya and Mt Meru
Apis mellifera sahariensis (the Saharan Honey Bee) – Moroccan desert oases of Northwest Africa, southern side of Atlas Range
Apis mellifera scutellata (the African Honey Bee) – widespread from South Africa to Somalia in eastern Africa. Its introduction to the Americas is reviewed by Winstonxxiii
Apis mellifera unicolor (the Madagascan Honey Bee) – Madagascar
The giant honey bees (subgenus Megapis)
There are three, perhaps four is one single extant giant honey bees, Apis dorsata – Indian subcontinent to Southeast Asia: China, Indonesia, India, Pakistan, and Sri Lanka, Philippines (Figure 10). There are four recognised geographically distinct subspecies:
Apis dorsata dorsata – widespead across most of southern Asiaprimarily India
Apis dorsata binghami – Sulawesi Malaysia, Indonesia
Apis dorsata breviligula – Philippines excluding western Palawan Island group
Apis dorsata laboriosa (Giant Mountain [Himalayan Cliff] Honey Bee) – Vietnam, Myanmar, Laos, southern China, eastern India (Nagaland, near Saramati Mountain)
Apis dorsata parasites include:
Acarapis dorsalis
Acarapis externus
Tropilaelaps clareae originated as a parasite of Apis dorsata laboriosa (Philippines and Sulawesi except Palawan Island, Luzon Island). It is now also a parasite of Apis cerana, Apis dorsata, Apis florea and Apis mellifera.
Tropilaelaps koenigerum is a parasite of Apis dorsata breviligula (Asia including Indonesia except the Sulawesi). It is now also a parasite of Apis dorsata,, Apis dorsata laboriosa and Apis mellifera.
Tropilaelaps mercedesae is a parasite of Apis dorsata laboriosa (Asia except Sulawesi including PNG). It is now also a parasite of Apis cerana, Apis dorsata and Apis mellifera.
Tropilaelaps thaii is a parasite of Apis dorsata laboriosa (Vietnam). It is now widely distributed in South and Southeast Asia and is found mainly in forested areas: Terai of Nepal, Malaysia and Singapore.
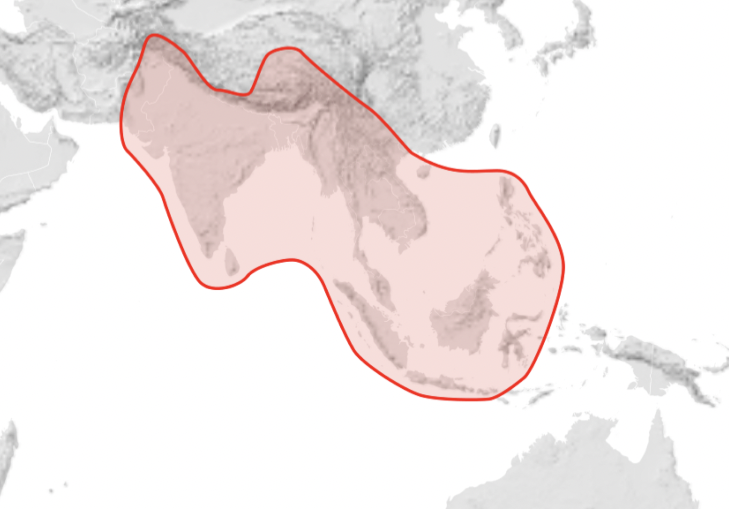
Figure 10 Apis dorsata distributionxxiv
Information on distribution of subspecies of Apis dorsata is sparse. However Apis dorsata laboriosa is very distinctive, is the largest of all living honey bees and its isolation and its adaptation to high altitude has sometimes accorded it separate species status, namely as Apis laboriosa.
Ancestral honey beesxxv
There are a number of extinct honey bee species known from the fossil record, one from the New World. These species fall into three subgenera, two unrelated to extant species groupings.
Fossil Apis species (subgenus Cascapis)
Apis armbrusteri – (Miocene) Germany Europe
Apis neartica – (Miocene) Nevada North America
Fossil Apis species (subgenus Megapis)
Apis lithothermaea – (Miocene) Japan
Fossil Apis species(subgenus Synapsis)
There are six well-documented extinct honey bees:
Apis cuenoti – (Oligocene) Europe
Apis henshawi (Henshaw’s Honey Bee) – (Oligocene) Creské Stredhori Mountains of Czech Republic, Europe
Apis longtibia (the Long-legged Honey Bee) – (Miocene) Shandong Province, China
Apis miocenca (the Chinese Miocene Honey Bee) – (Miocene) Shandong Province China
Apis petrefacter (the Petrified Honey Bee) – (Miocene) Creské Stredhori Mountains of Czech Republic, Europe
Apis vetustus (the Aged Honey Bee) – (Oligocene) Germany, Europe
Putative fossil Apis species
Other now extinct species have been cited in the literature, though their former existence is less certain:
Apis catanensis
Apis enigmatica
Apis trigona
Readings
iAddress for correspondence: 12 Lawrence Crescent, Kambah ACT 2902
awade@grapevine.com.au
iiSheppard, W. and Meixner, M. (2003). Apis mellifera pomonella, a new honey bee subspecies from Central Asia. Apidologie 34(4): 367-375. https://hal.archives-ouvertes.fr/hal-00891797/document
iiiWikipedia Honey bee https://en.wikipedia.org/wiki/Honey_bee
ivCervancia, C.R. (2002).Biodiversity of Apis cerana Fabricius in the Philippines. http://agris.fao.org/agris-search/search.do?recordID=PH2002000835
vArias, M.C,. Tingek, S., Kelitu, A.and Sheppard, W.S. (1996). Apidologie 27: 415-422. Apis nuluensis Tingek, Koeniger and Koeniger, 1996 and its genetic relationship with sympatric species inferred from DNA sequences. https://hal.archives-ouvertes.fr/hal-00891385/document
viArias M.C. and Sheppard, W.S. (2005). Phylogenetic relationships of honey bees (Hymenoptera:Apinae:Apini) inferred from nuclear and mitochondrial DNA sequence data. Molecular Phylogenetics and Evolution 37:25–35. https://www.ncbi.nlm.nih.gov/pubmed/16182149
viiiNotes on honey bee mites of the genus Acarapis hirst (Acari: Tarsonemidae) Delfinado-Baker, M. and Baker, E.W. (1982). International Journal of Acarology 8(4): 211-226. Published online: 17 Mar 2009 http://www.tandfonline.com/doi/abs/10.1080/01647958208683299?journalCode=taca20 [abstract only]
ixWikimedia Commons Apis florea distribution map.svg https://commons.wikimedia.org/w/index.php?curid=13706882
xhttps://commons.wikimedia.org/w/index.php?curid=13706485
xiKoetz, A. (2013). The Asian honey bee (Apis cerana) and its strains – with special focus on Apis cerana Java genotype – Literature review, Department of Agriculture, Fisheries and Forestry, State of Queensland, 59 pp. http://asianhoneybee.net.au/wordpress/wp-content/uploads/2013/03/AHB-behaviour-lit-review-FINAL2013.pdf
xiiEngel, M.S. (1999). The taxonomy of recent and fossil honey bees (Hymenoptera: Apidae: Apis). Journal of Hymenoptera Research 8: 165–196. https://kuscholarworks.ku.edu/bitstream/handle/1808/16476/Engel_JoHR_8%282%29165.pdf?sequence=1&isAllowed=y
xiiiSee explanatory note on the origins, evolution and species/subspecies association: Anderson, D.L. (2001). Varroa-bee relationships – what they tell us about controlling Varroa mites on the European Honey bee. https://www.apimondia.com/congresses/2001/Papers/396.pdf
xivRadloff, S.E., Hepburn, C., Hepburn, H.R., Fuchs, S., Hadisoesilo, S., Tan, K., Engel, M.S. and Kuznetsov, V. (2010). Population structure and classification of Apis cerana. Apidologie 41(6): 589-601. http://www.apidologie.org/articles/apido/full_html/2010/06/m08176/m08176.html at https://hal.archives-ouvertes.fr/hal-00892035/document
xvLo, N., Gloag, R.S., Anderson, D.L. and Oldroyd, B.P. (2010). A molecular phylogeny of the genus Apis suggests that the giant honey bee of the Philippines, A. breviligula Maa, and the Plains Honey Bee of southern India, A. indica Fabricius, are valid species. Systematic Entomology 35(2):226–233. The Royal Entomological Society. doi:10.1111/j.1365-3113.2009.00504.x
xviWikipedia Apis cerana nuluensis https://en.wikipedia.org/wiki/Apis_cerana_nuluensis
xviiWikipedia Apis nigrocincta https://en.m.wikipedia.org/wiki/Apis_nigrocincta and https://commons.wikimedia.org/w/index.php?curid=13709493
xviiiAnderson, D.L. R.B. Halliday, R.b. and and G.W. Otis G.W. (1997). The occurrence of Varroa underwoodi (Acarina: Varroidae) in Papua New Guinea and Indonesia. Apidologie 28: 143-147. https://hal.archives-ouvertes.fr/hal-00891413/document
xixWikipedia Apis koschevnikovi https://en.m.wikipedia.org/wiki/Apis_koschevnikovi
xxWhitfield, C.W., Behura, S.K., Berlocher, S.H., Clark, A.G. Johnston, J.S., Sheppard, W.S., Smith, D.R., Suarez, A.V., Weaver, D. and Tsutsui, N.D. (2006). Thrice out of Africa: ancient and recent expansions of the honey bee, Apis mellifera. Science. 314(5799): 642–645. doi:10.1126/science.1132772. PMID 17068261 Archived from the original (PDF) on September 29, 2015. http://www.life.illinois.edu/suarez/publications/Whitfield_etal2006Science.pdf
xxiRuttner, F., Tassencourt, and L. Louveaux, J. (1978). Biometrical statistical analysis of the geographic variability of Apis mellifera L.* I. Material and Methods. Apidologie 9(4): 363-381. https://hal.archives-ouvertes.fr/hal-00890475
xxiiCridland, J. M., Tsutsui, N. D. and Ramírez, S. R. (2017). The complex demographic history and evolutionary origin of the western honey bee, Apis mellifera. Genome Biology and Evolution 9(2):457-472. https://academic.oup.com/gbe/article/9/2/457/2970293/The-Complex-Demographic-History-and-Evolutionary?searchresult=1
xxiiiWinston, M.L. (1992) The biology and management of africanized honey bees. Annual Review of Entomology 37:173-93. http://www.annualreviews.org/doi/abs/10.1146/annurev.en.37.010192.001133?journalCode=ento
xxivWikimedia Commons Apis dorsata distribution map.svg https://commons.wikimedia.org/w/index.php?curid=13706771
xxvSee also reference to Engel loc cit; Hepburn, H.R. and Radloff, S.E. (eds) (2011). Honeybees of Asia. Springer -Verlag Berlin Heidelberg, , Germany. pp 207-214. https://books.google.com.au/books?id=zNuwxBP8MTUC&pg=PA47&lpg=PA47&dq=apis+miocene&source=bl&ots=u66tgaJsGU&sig=WZZOGV7gIks7XlsERAYKj_aAOT4&hl=en&sa=X&ved=0ahUKEwi6mr6566XRAhWFGJQKHZgTCOoQ6AEIFjAD#v=onepage&q=apis%20miocene&f=false
Engel, M.S., Hinojosa-Díaz, I.A. and Rasnitsyn, A.P. (2009).A honey bee from the Miocene of Nevada and the biogeography of Apis (Hymenoptera: Apidae: Apini). Proceedings of the California Academy of Sciences, Series 4, 60(3): 23–38. http://susquehannabeekeepers.com/pdfs/A_honey_bee_from_the_Miocene_of_Nevada_and_the_bio.pdf