

Demonstration of hive splitting: Tetragonula carbonaria colony at a Eurobodalla Botanical Gardens pollination workshop, New South Wales far south coast 12 October 2023:
(a) upper and lower spiral brood nest occupying separate brood chambers; and
(b) filled eke – honey super – that was positioned above a centrally placed baffle board atop brood chambers below.
Alan Wade
In examining the life of bees other than corbiculate bees we have drawn some conclusions about their approaches to their social lives. We now put each of the corbiculate tribes under the microscope, that is from the perspective of examining nests and members of each of the tribes, to see what they reveal.
Family Apidae: Apid bees
Sub family Apinae – Corbiculate bees
Having also traced the origins of the corbiculates, and having noted the extinction of a few sister tribes, we can now look more closely at their life systems. In attempting to define their social characteristics, we are limited by the fact that of the many species, particularly amongst the stingless and orchid bees, few have been well studied. Nevertheless we shall see that there are not only clear tribal differences but also that there are some striking variations in lifestyles even at the species level.
The nest
The corbiculate bee nest is best characterised as a community of bees occupying a secluded cavity with aggregated brood cells with stored provisions, features that even the orchid bees – that sometimes nest communally – possess. The cavity is occupied by a community of bees comprising a queen and often workers, where:
- in the case of honey bees, the colony is also seasonally accompanied by resident drones;
- in the case of stingless bees, the colony harbours virgin queens but does not retain drones;
- in the case of bumble bees, the colony produces numerous queens and drones as it goes into decline; and
- in the case of orchid bees – while lacking a worker class – colonies are sometimes multi queened, containing either unrelated queens or several generations of the foundress queen.
The queen
A defining feature of eusocial corbiculate bees is that their colonies are normally headed by a single queen. Amongst honey and stingless bees there may be additional gynes and very rarely a second mated queen – a makeshift short-lived arrangement – as contingency for queen replacement. The evolved condition of the eusocial corbiculates is that of workers ceding their reproductive capacity to a single queen upon which the colony is entirely dependent to build its workforce and to reproduce.
However there are a few exceptions. The stingless bees Melipona bicolor and Melipona quadrifasciata support more than a single queen.i
The drone
The role of the male corbiculate bee, the drone, is frequently misrepresented. Often characterised as the flying gamete – a misnomer when it comes to inbred diploid males – their role, taking into account the hymenopteran skewing consequence of haplodiploidy is no different to that of males of any species in the animal kingdom. However since the sperm of an individual drone are identical the queen he mates with produces closely related (super sister) daughters.
The important role of the corbiculate drone is well illustrated by the reproductive strategy of bumble bees. The frequent occurrence of queen loss in well established colonies serves a surprising purpose. Queenright bumble bee colonies produce massive numbers of well nurtured females, workers needed to build the colony and to produce queens to overwinter that will establish colonies anew in spring. In the normal functioning colonies the bumble bee queen effectively suppresses worker egg laying – as occurs also in honey and stingless bee colonies – and some drones are raised. Meanwhile bumble bee colonies that become queenless are stabilised by dominant workers and become an important source of drones to mate with queens produced by normal queenright colonies. Unsurprisingly we conclude that the true mission of all corbiculate bees is that of reproduction not to produce honey for humans and bears as apiarists (or bears and like predators) may assert. Honey and stingless bees store honey and pollen to enable the colony to expand and reproductively swarm and to tide itself over periods of dearth. On the other hand bumble bees invest their efforts in producing reproductives, foundress queens and drones to reestablish colonies in the following spring or early summer.
While honey bee drones remain with their parent nest or drift to other colonies, and remain dependent on them during maturation, other corbiculate drones leave the nest upon eclosure (emergence) and visit flowers for sustenance, though stingless bee drones appear to mature in a brief period before departing. Orchid bee drones are interesting in that they are sometimes associated with orchids, most famously as single species pollinators.
Both honey and stingless bees assemble in drone congregation areas, honey bees drones in well defined locations distant from honey bee nests while stingless bee drones create assemblages in close proximity to nests other than their own, zones that both virgin queens and drones alike seek. Bumble bees similarly mate beyond the confines of the hive but more in a one-to-one relationship, the queens then seeking out sheltered locations to go into diapause.
Drones in general appear to have no colony functional role in nurturing brood or harvesting for colony needs though they may be integral to honey bee colony homeostasis. Here we might return to the fact that within the Ceratinini tribe – the small carpenter bees – that one species, Ceratina nigrolabiata, employs male bees to protect nests.
Table 5.1 depicts the roles male (or drone) bees perform amongst the corbiculate tribes.
| Tribe | Mating location | Queen polyandry | Drone behaviour | Production timetable |
| Apini (honey bees) | Drone congregation areas (DCAs) remote from colonies | Multiple matings (number of matings species dependent) | Sustained by parent nest or drift to other nests; seek DCAs | Seasonal |
| Meliponini (stingless bees) | Drone congregation areas proximate to, but remote from, parent colonies | Mainly monandrous | Leave nest after a few weeks at most and rest on flowers or in communal roosts; seek DCAs | Continuous |
| Bombini (bumble bees) | Open mating | Monandrous | Abandon nest | Late summer or autumn |
| Euglossini (orchid bees) | Open mated | Monandrous | Abandon nest | Upon eclosure |
Table 5.1 Corbiculate drone mating behaviours.
The worker
Amongst the vast majority of insects, and indeed amongst nearly all hymenopterans, both males and females adopt a solitary existence. Inevitably, however, the burden of raising offspring falls entirely to the female of the species: a defining consequence of eusociality is that of formation of female castes and labour division. The colony generates a large number of workers that do ‘most of the heavy lifting’, not least – except for the labour of the queen laying eggs – in generating more of their own kind. At the same time the queen is protected by her offspring, is safely housed in a well secluded nest and averts – at least amongst honey and stingless bees – the hazards of foraging.
The primary roles of workers in all the social corbiculates are those of foraging, cell building, larval provisioning, nest debris removal and colony defence. However any notion of the worker being purely a subordinate member of the colony is a misnomer. They share or control many colony functions and are key to decision making, notably in respect of comb building, larval provisioning, swarming and harvesting of stores. At he same time queens assume the dominant reproductive roles of egg laying, suppressing worker egg laying, suppressing production of rival queens and exercising some control over drone production.
One of the more spectacular achievements of foraging workers of eusocial honey bees is their ability to communicate food and material resources. While the celebrated von Frisch discovery of dance signalling of flower resources is well known, less appreciated is the fact that honey bees remarkable detection of plant and other natural volatiles plays a major role in targeting flower and water resources.ii Spray lavender oil into a honey bee nest and bees will emerge in large numbers, fly across wind and head upwind to any patch of lavender in the disctrict. Stingless bees are different. Investigations to date have shown that the dance routine is entirely absent and that pheromone trailing is but one of many ways in which floral sources are indicated (Figure 5.1).iii Grüter canvasses many other ways by which these bees may communicate the finding of resources. However it is clear that eusocial bees in general have evolved many ways of signalling sources: food, water, propolis… to nest mates. This efficiency dividend may well explain why large colonies of bees gather far more honey per bee than do founding and weak colonies. Indeed the eastern Australian sugar bag bee, Tetragonula carbonaria, recruitment to food sources is enhanced if that source is upwind and scented.
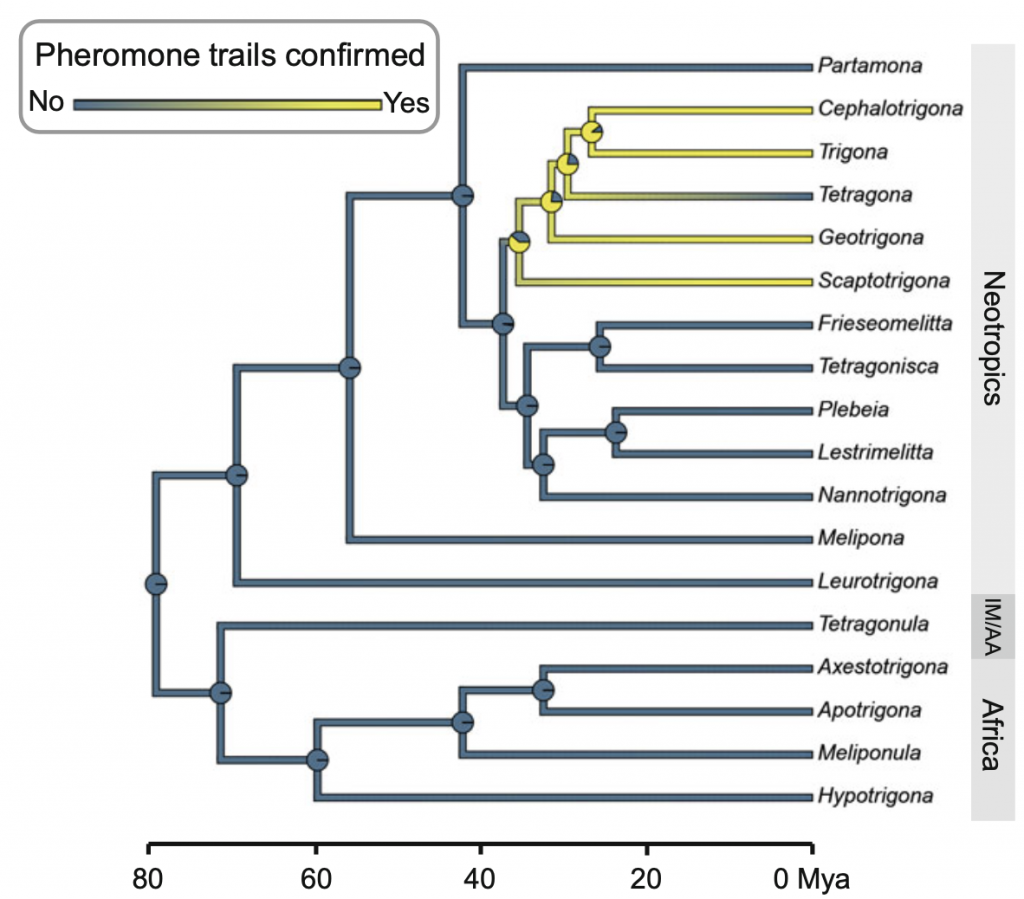
Figure 5.1 Frequency of pheromone trailing in stingless bees. The pheromone deposition trait was derived 30-35 mya.
The respective role of queens and workers amongst bumble bees is less well defined. Though only queens produce worker bees and potential queens, their reproductive dominance is less apparent. As we have noted strong colonies that become queenless can function normally though their role switches to that of drone production.
The self organising nature of corbiculate bee colonies, apart from the female skew in producing separate female castes, is a perhaps the only consistent emergent feature of their life systems. Let us look at the observable elements of each tribe and some of their signature species, the nest and all of its occupants, those many behaviours that have made their many social lifestyles so successful.
The Apini Tribe – Honey bees
The honey bee nest
The advanced western and asian honey bees nest in cavities, mainly tree hollows and rock crevices, the size choices of which have evolved to optimise overwintering survival and to optimise swarming frequency. Cavity dwelling honey bees have adapted to space-limiting nest walls by building multiple combs spaced to enable passage of bees and to facilitate ventilation. Several races have evolved to take advantage of the insulation and protection from predators afforded by shelter and radiated to cool temperate climes. With a propensity to hoard stores, some races of the asian honey bee Apis cerana (and the mountain adapted Apis nuluensis), and the western honey bee Apis mellifera have evolved to survive long winters and to the vicissitudes of elevated climes. Tropical races, as well as the open nesting dwarf and the giant honey bees migrate seasonally to follow the flowering pattern of plants while the giant mountain Cliff Honey Bee Apis laboriosa has adapted by seasonal altitudinal migration to aestivate in foothill forest floors.
Honey bee combs hang vertically and have two comb faces: their hexagonally fashioned cells are inclined slightly upwards from the horizontal (Figure 5.2). Not only does each cell share a wall with six other cells but the base of each cell is fashioned as a small inverted pyramid or pit walled by three interconnecting rhomboids, the faces of which are shared by three other cells on the opposing comb face.iv The cell geometry, not matched by other corbiculates, minimises cell building material use.
Comb architecture varies considerably across the honey bee genus. There is also considerable regional variation in cell size within a single species and hence worker progeny size.v David Heaf has observed that worker cell size varies considerably even within colonies, storage cells at the top of the comb being larger than brood cells formed on the lower comb margin.vi Worker, drone and queen cells are typically differentiated across the genus and both worker and drone comb are flexibly employed for brood raising and for storage of pollen and honey. The giant honey bees, however, produce larger cells for storage purposes and raise both workers and drones in the same sized cell.
The more primitive dwarf and giant honey bee clades nest on single combs attached to an overhang, often a tree branch or rock overhang, colonies often nesting in close proximity to one another and sited to achieve some protection from the elements.

Figure 5.2 Apis mellifera bees comb showing basic geometric features.vii
| Queen | Worker | Drone | ||||||||||
| Species | Egg | Larva | Pupa | Eclosure | Egg | Larva | Pupa | Eclosure | Egg | Larva | Pupa | Eclosure |
| Apis mellifera | 3 | 5 | 8 | 16 | 3 | 6 | 12 | 21 | 3 | 7 | 14 | 24 |
| Apis cerana | 3 | 4-5.5 | 6-7.5 | 13-16 | 3 | 5 | 11 | 19 | 3 | 6 | 14 | 23 |
| Apis dorsata | 3 | 4.5 | 7 | 14.5 | 2.9 | 4.6 | 10.9 | 18.4 | 2.9 | 4.6 | 14.3 | 21.8 |
| Apis florea | 3 | 6.8 | 7.7 | 17.5 | 3 | 6.3 | 11.2 | 20.5 | 3 | 6.7 | 12.8 | 22.5 |
Table 5.2 Brood development schedule in days for representative species of honey bee.viii
Honey bee larvae of all castes are capped at the 5th instar larval stage when they spin a cocoon, undergo metamorphosis and emerge, at the sixth and final stage, as adults. The pattern of development is the same across the Apis genus: queens eclose at about 15 days (16 d for Apis mellifera), workers around 20 days (21 d for mellifera) and drones rather later close to 23 days (24 d for mellifera). Notably the pupal stage for any particular caste hardly varies between species (typically 14 d). We may attribute the overall near invariant brood development period to close control of nest temperature (~35 0C), relative humidity (~ 80%) and to progressive provisioning. In a broodless state the cluster temperature is typically much lower (~18-20 0C in Apis mellifera).ix
The honey bee queen
While honey bees, like virtually all corbiculate bees, have evolved to be headed by a single queen, there are periods during queen succession where there may be a few to numerous gynes (potential queens) present. There is of course the common exception of finding a supersedure daughter queen laying alongside her parent.
Recently I came across such a colony with two queens (Figure 5.3). The bees had been rehived into a nucleus box from a wild nest in the autumn of 2023 and had survived the southern hemisphere winter. Our supposition was that the queen was old and, by late spring, had been superseded. The likely scenario was that the old queen had a low mandibular pheromone output and was not recognised by the new queen. Due to her age this queen would unlikely to have been a match for the new queen. However by very early summer, and just one week later, one of the queens had disappeared.

Queens wear out and need to be replaced periodically to ensure the long term survival of the colony. Typically established colonies of Apis mellifera persist for as long as 5-6 years,
but it is rare for a queen to live longer than 1-2 years. How honey bees handle queen loss or being headed by an underperforming queen depends on the species of bee and more particularly by the conditions under which a queen will be replaced. We note that:xi
- if a honey bee queen dies suddenly the bees will raise a new queen. The parent colony survives;
- if a queen is underperforming honey bee colonies have a simple remedy. The queen will be superseded, when the colony is very often headed by two queens, a mother and a daughter queen. The parent colony soon reverts to its ancestral single-queen condition;
- when a colony swarms, it takes with it the colony queen. However, if she is failing, the swarm will depart with a new queen and the old queen will be superseded. In either instance the parent colony is furnished with a new queen;
- if the colony continues to swarm, each new swarm will get one or more virgin queens. The parent colony again ends up with a new queen; and
- if the parent colony is over taken by an invading (parasitising) swarm its queen will be usurped.
In a remarkable insight into the differences between swarming and supersedure in honey bees, John Hogg noted that:xi
Queen supersedure is never the cause of swarming. But supersedure may occur concurrently with the swarm whenever the queen’s failure was caused by being over-taxed while generating the bees for that swarm. Significantly, it is the virgin queen that is then selected to accompany the swarm. The failed queen is retained in the parent hive to be superseded in turn…
However if requeening fails and a colony becomes queenless it will almost invariably dwindle and die. Apis florea colonies are well known to abscond rather than to raise a new queen so will automatically become a drone reservoir for as long as the colony survives. The Cape Honey Bee, Apis mellifera capensis, is similarly reluctant to raise an emergency queen. In a process called thelytokous parthenogensisxii it will very often rely on self requeening from unfertilised worker bees.
The most remarkable, and perhaps the most common, of these queen replacement strategies is swarming. It serves two purposes, colony increase and queen replacement. It involves colony fission where the old queen in the hollow log is replaced by a new queen (as also occurs in supersedure) and where the departing swarm is supplied either with the old queen (the primary swarm) or with one to many virgin queens (secondary swarms). Since the parent colony, or swarms accompanied by virgin queens, only allow the queen to depart on a mating flight once all rival gynes have been eliminated, both parent colony and swarms are always at risk of catastrophic queen failure. Wherever an old queen is involved, queen supersedure may also occur either after or as part of the swarming process.
The honey bee drone
Drones have extraordinarily well developed eyesight, notably for detecting sky (polarised UV and blue) light. They have exceptional spatial orientation and exceptional flight muscles and hence queen pursuit capability, their physical fitness being a measure of their mating success. They may also to have a role in maintaining elements of nest homeostasis and are anyway inimical to the thermal dynamics of the colony.
In general corbiculate bees invest a large proportion of their resources in producing drones and a more limited number of replacement queens. From an evolutionary perspective this should be seen as not only a contribution to their reproductive success but also of their capacity to adapt to ever shifting environmental conditions.
The mating behaviour of honey bee queens in drone congregation areas, mating-at-a-distance, minimises the chance of production of inbred diploid male brood.xiii Highly inbred stock found in queen breeding operations often enough results in colonies raising too few workers to be self maintaining. This is a condition that natural stock have evolved to avoid.
The honey bee worker
The honey bee worker exemplifies the self organising capacity of the honey bee colony and is characterised by aged based task assignment. Young bees clean cells and provision larvae. Middle aged bees construct comb, receive and process nectar and guard the colony entrance. In contrast, older bees have either a scouting role – locating and communicating floral resources and nesting sites – or are principally harvesters and control many facets of colony behaviour.
In honey bees reproductive swarming is precipitate and is orchestrated by colony workers. Swarming involves division of a colony in prime condition and usually under favourable seasonal conditions and is not dependent on availability of nesting sites. Colony workers cease provisioning the queen, make her flight ready, and orchestrate swarm departure. Usually the swarm settles briefly close to the hive while some workers scout for a new and sometimes distant nesting hollow. The swarm may also bivouac, as do giant honey bees. for a day or two before relocating.
While there is limited evidence that colony workers may scout for the nest sites in the days preceding planned swarming, the main searching is conducted on the temporary cluster of swarming bees. This honey bee swarming strategy is always risky as there is no assurance that honey bee swarms will find a nesting site while late season swarms are likely to have insufficient time to establish and furnish adequate stores for overwintering. However without the burden of establishing a new nest, honey bee colonies that have swarmed are normally well placed to rebuild for periods of dearth.
The Meliponini Tribe – Stingless bees
The stingless bee nest
Stingless bees are mainly though not always cavity dwellers.xiv Most species nest in either tree or natural subterranean hollows or in termite nests, the majority being confined to the tropics. Species that nest in the open are encased in a layered batumen to ward off the elements. All species raise brood in separate urn shaped or globular cells built from cerumen, a mixture of propolis and wax. Cells are not reused and honey and pollen are stored in separate pots located at the brood nest periphery.
Stingless bee comb building strategy is strikingly different to that adopted by honey bees. In a comb building study of stingless bees, including four very similar Australian species, Oldroyd and Pratt observed that eight species added single cells to existing comb in succession, while the remaining seven species added cells synchronously, that is to separate comb sections at the same time. Of these eight species built comb as aggregated clusters (mainly in the successive mode of addition), while one species constructed semi-comb (comb patches resembling piles of shattered car windscreen) and six species built regular sheet comb all but one synchronously. The sequencing of cell building and the type of comb built is species specific, different patterns of comb construction being common amongst many other stingless bees. To complicate matters a little, one species Pleibia poecilochroa from Brazilxv builds either a flat or spiral comb while the Australian temperate zone species Tetragonula carbonaria builds spiral combs only (Figure 5.4)

Figure 5.4 Split colony of Tetragonula carbonaria hive at Broulee on the warm temperate New South Wales south coast showing:
(a) single spiral comb lower nest; and
(b) upper nest located in an upper super.
The brood nest is surrounded by wax and resin multi layered involucrum (Figure 5.5) that serves to protect and insulate brood. Beyond these protective barriers are pollen and honey storage pots interspersing darkened used brood comb (top right).

Figure 5.5 Nest periphery detail of Tetragonula carbonaria: Basal section of spiral comb showing regular worker comb with raised and morphologically distinct sealed and unsealed drone comb (left).
The stingless bee queen
Stingless bee colonies, like those of honey bees, are headed by a single queen. However, unlike honey bee queens, stingless bee queens rarely mate more than once. This arrangement results in the progeny of stingless bee queen having workers that are all super sisters, their relatedness a consequence of hymenopteran drones having identical sperm. While the stingless bee virgin queen mates in a drone congregation area proximate to her nest, it appears that those associations are made up of drones from many colonies and that she always mates with a drone from another colony.
Stingless bee queens are very often larger than workers, although in some instances smaller queens are raised in worker sized cells. The processes by which females are destined to become workers or queens vary widely. In Tetragonula carbonaria, for example, the merging of two identical worker cells results in development of a queen.
The stingless bee drone
Stingless bee drones leave the hive shortly after emergence so their continued maintenance are not a burden on colony reserves. Since they are routinely raised with brood irrespective of the need of a colonies to replace their queens, they are produced at a routine cost to raising worker bees. Their regular availability ensures that virgin queens held by hives in the district can be mated to either replace a failed queen or to mate a queen supplied by colonies to establish a nearby swarm founding colony. Amongst some but not all species of Melipona, however, queens appear to be attracted to drone congregation areas (DCAs) in much the same way as honey bee queens are to Apis DCAs.
Swarming to new nesting sites is commonly very proximate to the parent nest, for example several nests can be established in the same termite mound as the parent colony. This ensures ready transfer of hive materials and stores to the fledgling nest. Drones, comprised almost entirely of progeny from distant nests, congregate close to hives superseding their queen and to new colonies that the parent colony has supplied a virgin queen. Stingless bee drones have been observed to roost together and to visit flowers for sustenance.
The stingless bee worker
Very remarkably stingless bee workers may, in some cases, be active reproductives joining the queen in laying drone eggs. Christoph Grüter reports twenty two species where workers lay eggs in the presence of a queen, three species that lay in queenless colonies and seven species that never lay eggs. Of these some are produced as a food source, so-called trophic eggs, that are nutritionally rich but lack genetic material to develop, while other eggs become drones. The reproductive biology of stingless bees is extremely complex and variable, in part attributable to cell mass provisioning where an egg is laid onto a food bed when the cell is promptly sealed.
As a rule of thumb stingless bees swarm less regularly thatn do honey bees but the success rate of new colony establishment is higher. Lifespans of stingless bee colonies are variable and little understood though Grüter in surveying published records instances some colonies surviving for very many decades.
Stingless bee worker roles are those of most eusocial bees, undertaking the chores of nest selection, comb building, brood provisioning and harvesting. As with honey bees they scout for new nesting sites and initiate swarming.
Engles and Imperatriz-Fonseca provide a generalised account of stingless bee swarming process (Figure 5.6). In their model, scout bees locate a suitable nesting cavity close to the parent nest. Having ascertained that the cavity is suitable, they first bring cerumen – a mixture of plant resins or propolis and waxy bee secretions – to construct a protectable entrance tube and line the future nest with batumen (a mixture of soil and bee excrement and cerumen), construct storage pots and supply the fledgling nest with stores. To this they direct a swarm with a virgin queen from the parent colony to occupy the new nest. The new queen is mated with a drone proximate to the new nest. The nest is then well supported by the parent nest so the risk of its not being established is minimal but its establishment process is slow. The parent colony also furnishes a replacement queen in the event of the daughter colony losing its queen. However, dispersal of stingless bees (at least those in tropical jungles) is limited and colony densities may be relatively high if there are many potential nesting sites as has been observed in species nesting in termite mounds.
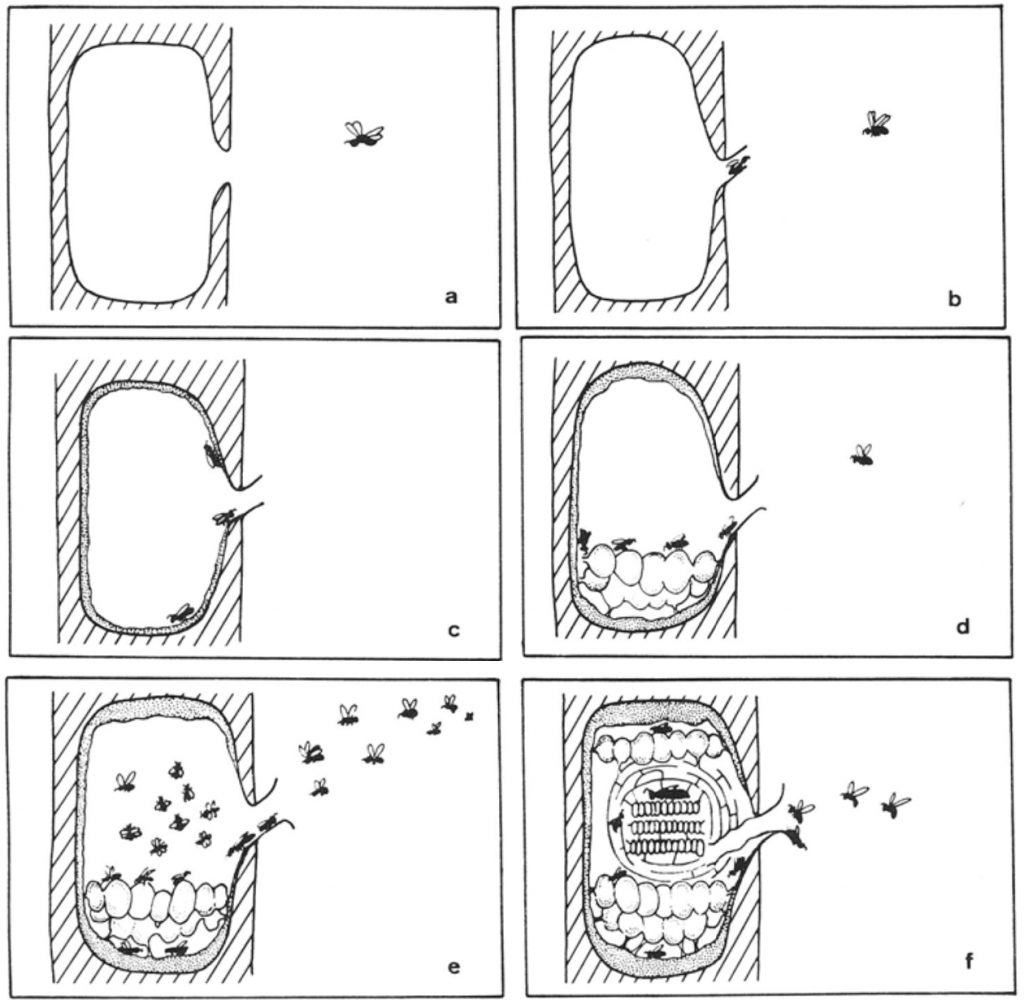
Figure 5.6 Stingless bee swarm colony establishment.xvi
The process of colony fission is sometimes extended to a period of months, the new colony becoming self sustaining dependent on many factors including the long period it takes for the new colony to raise its own workers.
I were once able to witness the similar budding off process that ants sometimes employ as a means of establishing new colonies. In studying the foraging behaviour of a nest of the meat ant, Iridomyrmex purpureus viridiaeneus in arid zone environment of the Kinchega National Parkxvii we discovered that ants forced to retreat from foraging in the eye of an approaching thunderstorm did not all return to the large mother nest. Some ants headed for a small nest some fifty or so metres distant. We surmised that this was a satellite nest, a daughter colony (Figure 5.7).
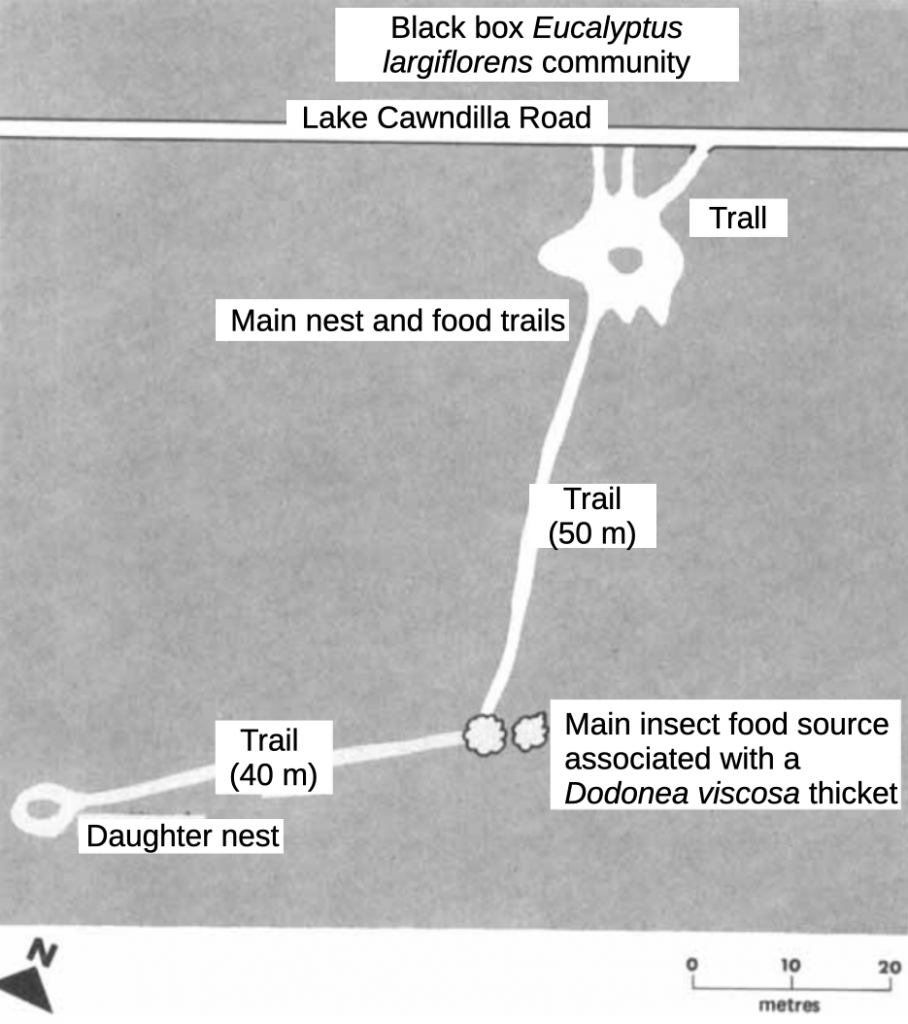
Figure 5.7 Iridomyrmex purpureus form viridiaeneus (meat ant) colony fission.
This self requeening strategy has a fundamental implication for the management of stingless bee hives. Strong colonies can be split judiciously (see Figure 5.4 and frontispiece), each section of the hive nadired (undersupered) to allow each split to rebuild its nest as if it had been allowed to swarm naturally. No new queen is needed as both have latent virgin queens.
The Bombini Tribe – Bumble bees
In reviewing the overall social status of bumble bees in the context of other corbiculate bees, Woodard and coworkers observed that:xviii
Eusocial insects live in groups with three defining characteristics: a reproductive division of labour, cooperative brood care and overlapping generations. As opposed to highly eusocial species, such as honey bees, which live in large perennial colonies, most bumble bees live a so-called ‘primitively’ eusocial lifestyle wherein queens are solitary for part of their annual colony cycle. The remainder of the colony cycle comprises a social group with a single reproductive queen and her typically non reproductive daughters until late in the colony life cycle, when new reproductive offspring are produced.
The bumble bee nest
Ah, look on my grievous woe; ah, would that I could turn
To a humming-bee, and so win through to thy shy retreat,
Lightly thridding the clinging ivy and sheltering fern.
The Idylls of Theocritusxix
Unlike stingless and honey bees, bumble bees nests are started anew each year and their populations are built from a single founding queen. In a study of bumble bees in southern England, Alfordxx found that young queens enter diapause for periods for as long as six to nine months signalling that the biology of overwintering queens is not incidental to their life cycle. Daughter queens often aestivate near the parent nest where nesting conditions tend to be favourable and where they may inherit the parent nest. Overwintering cuckoo bumble bee queens (subgenus Psithyrus) may also overwinter in the same vicinity but delay eclosure in order to usurp regular bumble bee queens in colonies once well established.
Michenerxxi outlines the nest founding process. Drawing heavily on studies of Alford,xxii and a much earlier study by Sladen,xxiii he outlines how overwintered queens first locate a nesting site, typically an abandoned rodent nest, a grass tuft, a bird box or amongst litter under trees. Sladen’s findings, updated by John Free and Colin Butler, provide more detail of their life systems.xxiv
To start a new colony the queen first forages and lines her nest, typically a small cavity 3-4 cm in diameter, where she deposits a pile of pollen she has collected either on the floor of the nest or in a just started cell. Onto this she lays an egg horizontally and subsequently, after more provisioning, a number of eggs before sealing each cell and moving on to construct an entirely new cell. Sladen describes how the queen settles over a central groove in the brood capsule to incubate larvae (Figure 5.8, schematically Figure 5.9) leaving the nest only to recharge adjacent honey pots that she will fill early each day to continuously sustain her brood. This central groove facility is lost as colonies expand and the nest becomes warmer.
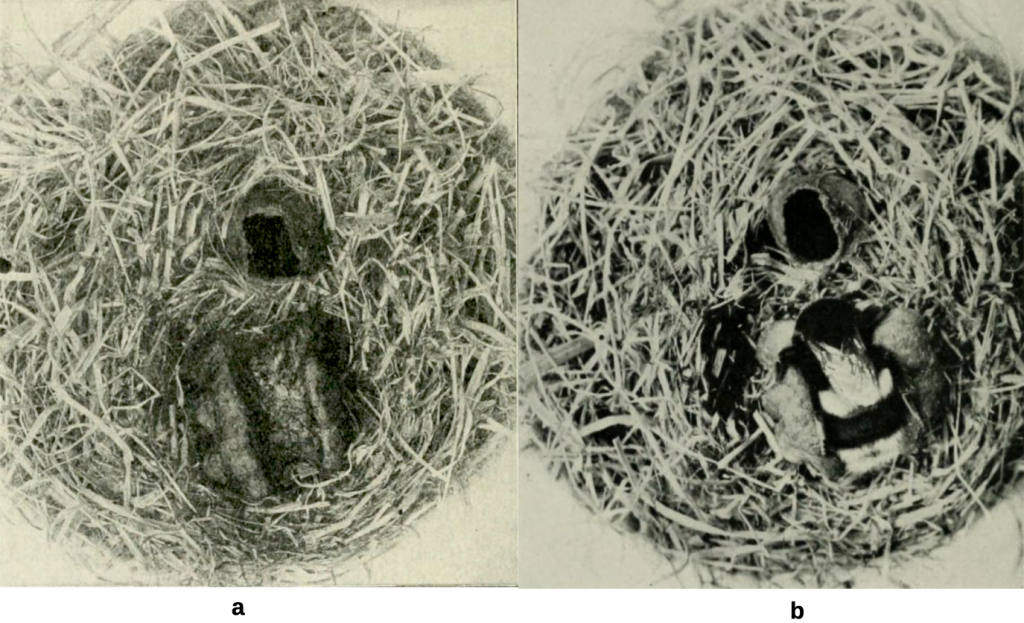
Figure 5.8 Bombus terrestris queen incubation of brood after Sladen:
(a) groove in surface of brood capsule to facilitate heat transfer; and
(b) queen nestled over brood that she feeds intermittently with a mixture of stored honey and pollen that she injects through a small hole in the waxen nest capsule.
As the colony develops the queen ceases foraging but, apart from her primary function as the egg layer, she continues in her role of provisioning brood, one that she shares with workers. Hence the respective roles of bumble bee worker and queen are less clearly delineated than they are amongst honey and stingless bees.
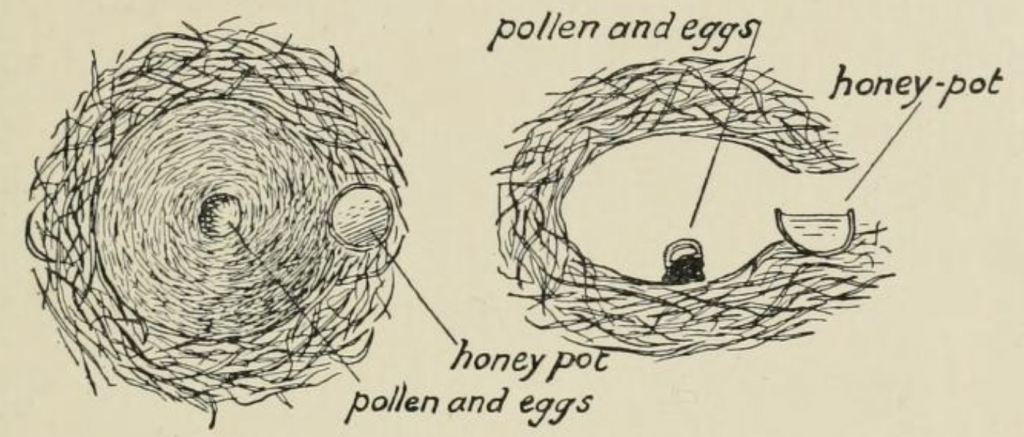
Figure 5.9 Nest initiation in bumble bees after Sladen.
Brood of bumble bees appear to be reared on much the same timetable as honey bees (Table 5.3), but development times vary considerably between species and even within the one nest. Whereas amongst honey bees only older workers become foragers, bumble bees may forage as early as the third day after emergence. The colony life cycle also varies widely being very short amongst more northerly species of bumble bees with some perenniality being found with tropical species.
| Taxa | Egg | Larva | Pupa | Eclosure |
| Bombus lapidarius/ Bombus hortorum | 4 | 7 | 11-12 | 22-23 |
| Apis mellifera | 3 | 6 | 12 | 21 |
Table 5.3 Worker brood development schedule comparing the bumble bees Bombus lapidarius and Bombus hortorum with the honey bee Apis mellifera.xxv
Notes: Bombus bee eggs are larger and more transparent than those of the common honey bee. Eggs and larvae (7-16 depending on species) in each capsule are covered with a waxen envelope that the queen enlarges as larvae grow. Cocoons are spun and stand erect side to side. Upon eclosure the queen or workers assist the larva by chewing out cocoon caps.
If the amount of food is suboptimal or if the weather is adverse bumble bee development may be extended up to a period of about one month.
Sladen also details differences in morphology and behaviour amongst bumble bees, especially those of the usurping subgenera Psithyrus species. While most bumble bee queens have well developed corbiculae, they are degenerate in honey and stingless bee queens, and also in parasitic Psithyrus bumble bees.
Eventually the queen is joined by emerging daughter workers assuming foraging and nest duties constructing an irregular clump (or clumps) of cells in the process raising more workers when the colony becomes eusocial. Food pots in various configurations are established on brood cell margins to ensure continuous brood provision and as contingency for periods of dearth. Two types food storage exist, the more primitive pollen pocket type (Figure 5.10a) and those with well defined storage pots (Figure 5.10b). A fairly typical nest (Figure 5.11) illustrates this complex multi cell assemblage. Details of the operation of these two nest types are expanded upon by Free and Butler, notably in respect of intermediate behaviour of Bombus impatiens and Bombus pratorum.xxvi
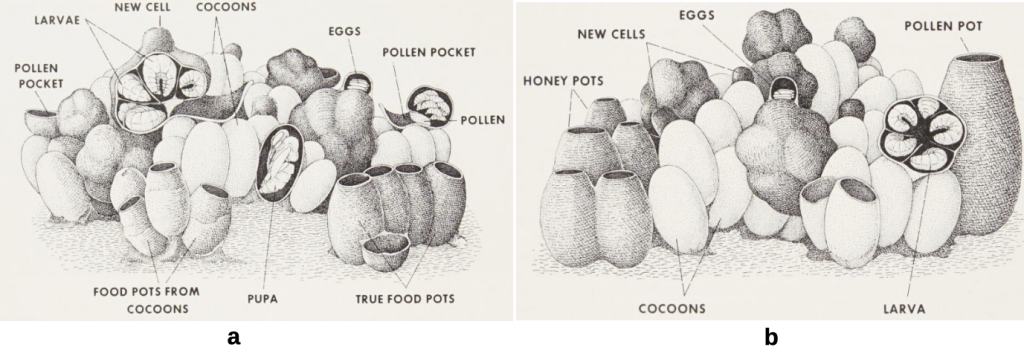
Figure 5.10 Brood nests of different bumble bees clades:
(a) a pocket making species Bombus (Fevidobombus) atratus; and
(b) a pollen storing species of Bombus.
Images from de Camargoxxvii redrawn by Michener.

Figure 5.11 The bumble bee nest.xxviii
The bumble bee queen
While colonies are established by overwintering foundress queens the chances of her successfully doing so are not high but some at least succeed. The trade-off in energy terms is, however, favourable as an overwintering colony of honey bees must contend with long periods of dearth, and is always at risk of running out of stores and starving as occurs with many late season honey bee swarms.
Having established a new nest, workers join and assist the queen in building a sizeable community, one that may expand to as many as several hundred individuals.
While early loss of a bumble bee queen in the nest development phase results in its demise, in well established bumble bee colonies the situation is very different to that of honey and stingless bees. Dominant workers assume a queenlike role laying drone eggs while the majority of workers continue to focus on comb building, on foraging and on brood nurturing. Though without a queen to bolster colony numbers, the colony nevertheless remains cohesive and becomes an important source of drones needed to mate with the many queens produced by queenright colonies.
The interplay of queen succession across the corbiculate tribes is a complex process but might be best summarised as ‘Where there is a will, there is a way’. The fact that all three tribes produce either swarms or foundress queens in relative abundance means that occasional loss of permanent colonies or loss of of aestivating queens is of little consequence. Interestingly not only is eusocial behaviour lost in colonies that become queenless, but is also lost when bumble bee colonies sometimes evict their queen as the colony dwindles and dies.
In late summer or autumn queenright bumble bee colonies switch to producing larger cells in which they raise queens. These will mate with drones raised in both queenright and queenless colonies, mated queens then entering diapause to overwinter to begin the cycle again. Though many overwintered queens fail to establish new colonies, and some of those that do establish new colonies are parasitised, the investment in the production of many queens ensures species survival. This said many bumble bee species are in decline, habitat destruction and consequent inbreeding being cited as among the likely contributing factors.xxix
Temporarily eusocial bumble bees have a queen management strategy that is defined by a very different rules to those adopted by the honey and stingless bees: their queens are not normally replaced though now and then a queen may be usurped, a phenomenon also reported to also occur amongst honey bees.xxx Bumble bee colonies focus is on colony succession, that is annual production of large numbers of queens and drones, rather than on any form of queen replacement or colony persistence.
The bumble bee drone
Like stingless bee drones, and unlike honey bee drones, those of the bumble bee leave the nest shortly after emerging. Sladen describes how queens, released from the parent colony late in the season, mate with drones before they enter diapause:xxxi
Fertilisation is the first important event in the queen’s life. This takes place in the open air as a rule, but there are good reasons for supposing that it can be accomplished within the nest. The young queens are shy and show themselves very little. The males course up and down hedgerows, or hover over the surface of fields and around trees, in the hope of finding their mates. Each species has its particular kind of hunting-ground.
In this sense at least queens and drones behave very much as do solitary bees. After mating the female bee is left with all the tasks of raising new offspring.
The bumble bee worker
The supposition that the humble-bee worker is less industrious than the honey-bee is erroneous: she labours with the same zeal and tireless energy, never ceasing until, worn out, she fails one day to return home, and, becoming drowsy and senseless, passes out of existence in the cold of the succeeding night.
Sladen, 1912
As we have seen the founding bumble bee queen is the original worker. She undertakes all the duties of nest establishment, foraging and brood provisioning, that is until her early brood emerge to assist her. It is only then that the colony becomes eusocial. Unlike the facultatively eusocial bees found in the halictine sweat bee tribe (Part II) and the xylocopine carpenter bee tribe (Part III), bumble bees have no choice other than to enter the eusocial contract. They are simply not programmed to survive without the assistance of worker bees.
Having said this, the roles of workers and queens amongst bumble bees are not as well delineated as they are amongst honey and stingless bees. A strong colony that becomes queenless results in dominant workers taking over the role of egg laying – they only produce drones – and in maintaining worker roles of foraging, comb building and brood provisioning. The colony becomes sub social consisting of individuals of the same generation and necessarily becomes incapable of generating new queens and workers. From here the main workforce of both queenright and queenless colonies continue to undertake the normal colony roles of foraging, cell construction and brood provisioning.
As the season progresses, and as colonies mature, bumble bees turn to producing reproductives rather than workers. Instead of building stores and down regulating their populations to survive dearth, as do stingless and overwintering honey bees, bumble bees adopt the solitary bee strategy of instead raising queens that will overwinter. Bumble bee colonies, at least those with the prospect of winter, perish but face the near insurmountable task of establishing colonies from scratch each spring.
The Euglossini – Orchid bees and
Extinct Tribes – Lost eusocial bees
The orchid bee nest
Michener, in a review of the predominantly central American orchid bees, signals their close alliance to bumble bees in their starting solitary reproductive condition noting also the common occurrence of multi queen colonies.xxxii, xxxiii, xxxiv The arrangement of cells in both solitary and communal nests is clustered (Figure 5.12) though a linear cell arrangement sometimes occurs. Each cell is mass provisioned, as are stingless bees, and laid into by the queen in sequence.
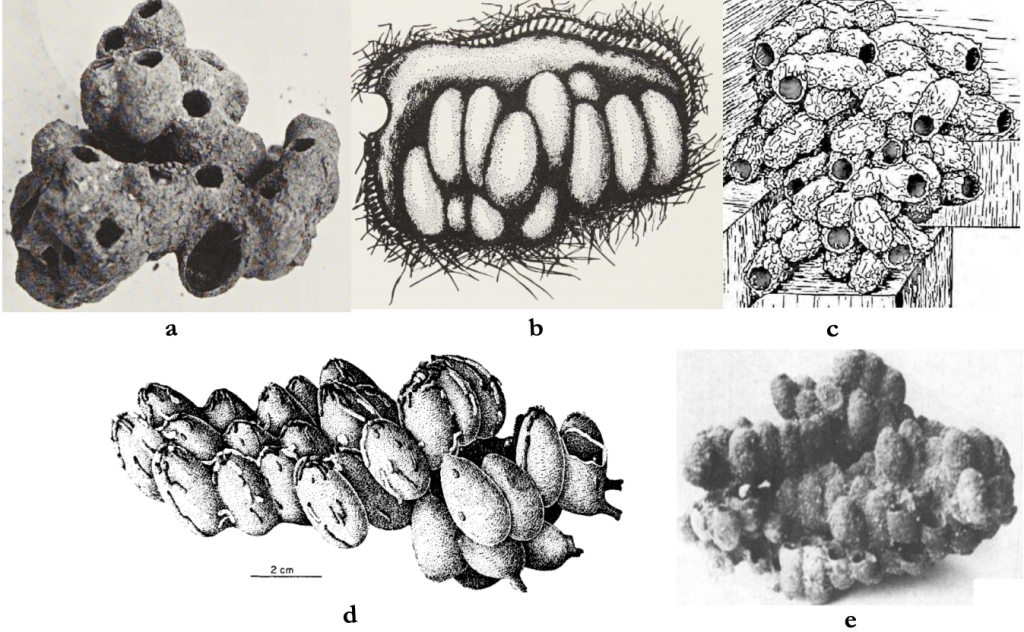
Figure 5.12 Orchid bee nest cell clustering:
(a) Eulaema terminata;
(b) Euglossa ignita;
(c) Eufriesia surinamensis;
(d) Eulaema nigrita; and
(e) Euglossa impcrialis.
Drawing on the findings of Zucchi, Sakagami and de Camargo,xxxv Michener attempts to clarify the social status of multi queened orchid bee colonies. They focused their study of the nesting behaviour of species of Euglossa and Eulaema. Lone females establish their own nest, where subsequently some daughters leave the nest while others stay on to form either separate or merging clumps of cells. Their daughters, in turn, may mate and remain to form yet more nests resulting in a colony with a massive cluster of cells and a large number of female bees.
That more adults may be present than cells being built or larvae present suggests that some Euglossini may be quasi social. Again citing Zucchi, Sakagami and de Camargo, Michener refers to a specific study of a nest Eulaema nigrita with three females. Each female constructed and provisioned her own nest. However resident females that were observed to assist with nest duties (such as cell construction) were prevented from doing so by the brood owner returning from foraging.
So while there may be some joint provisioning of larvae in orchid bee colonies, there is no defined worker caste, all females being mated and raising their own brood. Euglossini bees, though typically solitary, may alternatively be highly social but never eusocial.xxxvi
The orchid bee queen
Orchid bees may nest communally as multi queened colonies and have also been occasionally reported to usurp the larval stores of females who fail to return to the nest..xxxvii The general consensus is that orchid bees are solitary in reproductive terms – no female skew is evident – and that some taxa have strong social traits. However since orchid bees lack a worker caste whether one assigns the term queen rather than female is perhaps purely semantic.
The orchid bee drone
The males of some orchid bee species are well known to be orchid pollinators, hence their epithet. In general they leave the nest, as also do most females, upon eclosure (emergence). Their role as pollinators appears to be commensal since, as well as depending on flowers for sustenance and acting as pollinators, they also appear to collect the scent from flowers to make themselves attractive to female orchid bees.xxxviii
The working orchid bee and extinct corbiculate bee tribes
The working orchid bee is, of course, the solitary queen (or female) as she also builds the nest and provisions her offspring even in a communal setting.
That orchid bees have reverted in the main from an ancestral eusocial condition to a purely solitary condition is a good point to exit the discursive topic of the social life of bees. One wonders what Euglossopteryx biesmeijeri (Figure 5.13), an extinct solitary member of an incertae sedis (i.e. an unassigned corbiculate tribe), or what the other known extinct eusocial corbiculate tribes, Melikertini, Electrapini, and Electrobombini, might have thrown up.xxxix Sufficient to say is that surviving corbiculate tribes have clearly benefited from a cooperative, diverse and mainly eusocial way of life.

Figure 5.13 Fossil Euglossopteryx biesmeijeri from Eocene oil shale deposit, Green River Formation, Uinta Basin, Utah.
Readings
iGrwswüter, C. (2020). Stingless bees: Their behaviour, ecology and evolution, 395pp. Fascinating Life Sciences. pp.151-152. https://sci-hub.mksa.top/10.1007/978-3-030-60090-7
iiTautz, J. (2023). Honey bee communication – a revision. The Australasian Beekeeper 125(1):49-50.
Tautz, J. (December 2022). The new three-phase model in honey bee recruiting. Bee World 99(4):1-2. doi:10.1080/0005772X.2022.2151743
Tautz, J. and Steen, D. (2017). The honey factory: Inside the ingenious world of bees. The waggle dance: Rethinking old concepts, pp.67-111. Black Ink, Carlton, Victoria.
iiiGrüter (2020) loc. cit. Chapter 10 Recruitment and communication in foraging. pp.340-366.
ivBauer, D. and Bienefeld, K. (2013). Hexagonal comb cells of honeybees are not produced via a liquid equilibrium process. Naturwissenschaften [The Science of Nature] 100(1):45-49. doi:10.1007/s00114-012-0992-3 https://www.researchgate.net/publication/233411536_Hexagonal_comb_cells_of_honeybees_are_not_produced_via_a_liquid_equilibrium_process
vWade, A. (2023). The honey bee comb catacomb. The Beekeepers Quarterly 151:38-42.
viHeaf, D. (2010). The bee-friendly beekeeper: A sustainable approach. pp.41-42. Northern Bee Books, Scout Bottom Farm, Mytholmroyd, West Yorkshire.
viiChamberland, M. (22 July 2015). The miraculous space efficiency of honeycomb: Hexagons and the science of packing. https://compote.slate.com/images/7e013162-4951-4a8c-927c-404274b3ec21.jpg
viiiOldroyd, B.P. and Wongsiri, S. (2009). Asian honey bees: Biology, conservation, and human interactions. Harvard University Press. http://trove.nla.gov.au/version/45924294
ixKastberger, G., Waddoup, D., Weihmann, F. and Hoetz, T. (2016). Evidence for ventilation through collective respiratory movements in giant honeybee (Apis dorsata) nests. PLoS One 11(8):e0157882.
doi:10.1371/ journal.pone.0157882
Tan, K., Yang, S., Wang, Z-W., Radloff, S. and Oldroyd, B. (2012). Differences in foraging and broodnest temperature in the honey bees Apis cerana and A. mellifera. Apidologie 43(6):618-623. https://hal.science/hal-01003658/file/hal-01003658.pdf https://www.tib.eu/en/search/id/BLSE%3ARN321836196/Differences-in-foraging- and-broodnest-temperature/
xSeeley, T.D. (2017). Life-history traits of wild honey bee colonies living in forests around Ithaca, NY, USA. Apidologie 48(6):743–754. https://link.springer.com/article/10.1007/s13592-017-0519-1 doi:10.1007/s13592-017-0519-1
xiHogg, J.A. (2006). Colony level honey bee production: The anatomy of reproductive swarming. American Bee Journal 146(2):131-135. http://www.twilightmd.com/Samples/Hogg/Hogg_Halfcomb___Publications/ABJ_2006_February.pdf
xiiEsko Suomalainan describes three types of parthenogensis: Arrhenotoky where unfertilised eggs develop parthenogenetically into males; Thelytoky where unfertilised eggs develop into females; and Deuterotoky or Amphitoky where unfertilsed eggs develop into both sexes. Suomalainen, E. (1950). p.196. Parthenogenesis in animals. Advances in Genetics 3:193-253. doi:10.1016/S0065-2660(08)60086-3
xiiiCollison, C. (2023). Queen/drone mating: A closer look. The Australasian Beekeeper 125(3):50-52.
xivRoubik, D.W. (2020). Nest structure: Stingless bees. In Starr, C.K. (ed). Encyclopedia of Social Insects. pp.646-651. doi:10.1007/978-3-319-90306-4_147-1 https://sci-hub.mksa.top/https://sci-hub.mksa.top/10.1007/978-3-030-28102-1_117
xvDrummond, P.M., Bego, L.R. and Melo, G.A.R. (1995). Nest architecture of the stingless bee Plebeia poecilochroa Moure & Camargo, 1993 and related considerations (Hymenoptera, Apidae, Meliponinae), Iheringia 79:39-45. https://www.biodiversitylibrary.org/page/34487305#page/749/mode/1up
xviEngels, W. and Imperatriz-Fonseca, V.L. (1990). Chapter 7. Caste development, reproductive strategies, and control of fertility in honey bees and stingless bees. In Engels, W. (ed). Social insects: An evolutionary approach to castes and reproduction. p.205. Springer, Berlin, Heidelberg. https://sci-hub.se/10.1007/978-3-642-74490-7_9 https://link.springer.com/chapter/10.1007/978-3-642-74490-7_9
xviiMobbs, C.J., Tedder, G., Wade, A.M. and Williams, R. (1975).A note on food and foraging in relation to temperature in the meat ant Iridomyrmex purpureus form viridiaeneus. Australian Journal of Entomology 17(3):193-197. doi:10.1111/j.1440-6055.1978.tb00142.x https://onlinelibrary.wiley.com/doi/pdf/10.1111/j.1440-6055.1978.tb00142.x
xviiiWoodard, S.H., Lozier, J.D., Goulson, D., Williams, P.H., Strange, J.P. and Jha, S. (2015). Molecular tools and bumble bees: Revealing hidden details of ecology and evolution in a model system. Molecular Ecology, 24(12):2916-2936. doi:10.1111/mec.13198
xixHallard, J.H. (1894). The Idylls of Theocritus. London, Longmans, Green and Co.. Idyll III. The desperate lover. p.15. https://ia600209.us.archive.org/1/items/cu31924026600324/cu31924026600324.pdf
xxAlford, D.V. (1969). A study of the hibernation of bumblebees (Hymenoptera: Bombidae) in southern England. Journal of Animal Ecology 38910:149-170. doi:10.2307/2743
xxiMichener, C.D. (1974). The social behavior of the bees: A comparative study. Harvard University Press. Bumble bees. Part III. Chapter 28. pp.314-328. https://archive.org/details/socialbehaviorof0000mich/page/314/mode/1up
xxiiAlford, D.V. (1970). The incipient stages of development of bumble bee colonies. Insectes Sociaux 17(1):1-10. doi:10.1007/bf02223768 https://link.springer.com/article/10.1007/BF02223768
xxiiiSladen, F.W.L. (1912). The humble-bee, its life-history and how to domesticate it with descriptions of all the British species of Bombus and Psithyrus. Chapter II. pp.1-35. MacMillan and Co. Limited, London, Bombay, Calcutta, Melbourne. https://archive.org/details/humblebeeitslife00slad
xxivFree, J.B. and Butler, C.G. (1959). Bumblebees. The New Naturalist volume 14. 208pp. Collins St Johns Place, London https://archive.org/details/bumblebees0000john_n2u2/page/n6/mode/1up
xxvOldroyd and Wongsiri (2009) loc. cit.
xxviFree and Butler (1959) loc cit. p. 17.
xxviide Camargo, J.M.F. (1990). Ninhos e biología de algumas espécies de Meliponideos (Hymenoptera: Apidae) da região de Pôrto Velho, Território de Rondonia, Brasil. Revista de Biología Tropical 16(2):207-239. https://tropicalstudies.org/rbt/attachments/volumes/vol16-2/04-Camargo-Meliponideos.pdf
xxviiiMiksha, R. (10 July 2015). Plight of the bumble bees. Bad Beekeeping Blog. https://badbeekeepingblog.com/2015/07/10/plight-of-the-bumblebees/
xxixWoodard, Lozier, Goulson, Williams, Strange and Jha (2015) loc.cit. pp.2196-2197.
xxxMangum, W.A. (May 1, 2009). Usurpation: A colony taken over by a foreign swarm. American Bee Journal 149(5):excerpt. https://americanbeejournal. com/usurpation-a-colony-taken-over-by-a-foreign-swarm/
Mangum, W.A. (2010). Honey bee biology: The usurpation (takeover) of established colonies by summer swarms in Virginia. American Bee Journal 150(12):1139-1144. Cited by Oliver, R. (2014). What’s happening to the bees? – Part 2. http://scientificbeekeeping.com/whats-happening-to- thebees-part-2/
Mangum, W.A. (2012). Colony takeovers (usurpations) by summer swarms: They chose poorly. American Bee Journal 153(1):73-75. Cited by Oliver, R. (2014) loc. cit.
Mangum, W.A. (2013) Summer swarms with queen balling. American Bee Journal 153(2):163-165. Cited by Oliver, R. (2014) loc. cit.
Mangum, W.A. (January 1, 2018). More usurpation: Colony take over by summer swarms. American Bee Journal 158(1):excerpt. https:// americanbeejournal.com/usurpation-colony-take-summer-swarms/
Mangum, W.A. (May 1, 2019). Usurpation: A colony taken over by a foreign swarm. American Bee Journal 159(5):537-541. https:// americanbeejournal.com/usurpation-a-colony-taken-over-by-a-foreign- swarm/ https://bluetoad.com/publication/?i=580808&article_ id=3356173&view=articleBrowser
Mangum, W. (December 1, 2020). Summer swarms: A prelude to usurpation (colony takeover). American Bee Journal 160(12):1321-1326. https://abj-fe8.kxcdn.com/wp-content/uploads/2021/01/abj_ december_2020_final-bt-sm.pdf https://americanbeejournal.com/summer-swarms-a-prelude-to- usurpation-colony-takeover/
Mangum, W.A. (December 1, 2021). Colony usurpation (takeover):
The fourth method of queen replacement – Part 1 American Bee Journal 161(12)::excerpt.https://americanbeejournal.com/colony-usurpation-takeover-the-fourth-method- of-queen-replacement-part-1/
Mangum, W.A. (January 1, 2022). American Bee Journal 162(1):excerpt.Colony usurpation (takeover): Situations that stall the takeover process – Part 2. https:// americanbeejournal.com/colony-usurpation-takeover-situations-that-stall- the-takeover-process-part-2/
xxxiSladen (1912) loc. cit. Chapter II. p.12.
xxxiiMichener, C.D. (1974). The social behavior of the bees: A comparative study. Harvard University Press. Part III, Chapter 21 The orchid bees (Euglossini). pp.257-260. https://archive.org/details/socialbehaviorof0000mich/page/257/mode/1up
xxxiiiKimsey, L.S. (1982). Systematics of bees of the genus Eufriesia. University of California Publications on Entomology 95:1-125. https://www.researchgate.net/publication/344288052_Systematics_of_Eufriesea_Kimsey_reducedsized
xxxivZucchi, R., Sakagami, S.F. and de Camargo, J.M.F. (1969). Biological observations on a neotropical parasocial bee, Eulaema nigrita, with a review on the biology of Euglossinae (Hymenoptera, Apidae): A comparative study. Journal of the Faculty of Science, Hokkaido University 17(2):271-380. https://www.semanticscholar.org/paper/Biological-Observations-on-a-Neotropical-Parasocial-Zucchi-Sakagami/a7f5b7be79873e368e817fdb7d9f049cec645583
xxxvZucchi, Sakagami and de Camargo (1969) loc. cit.
xxxviDodson, C.H. (1966). Ethology of some bees in the tribe Euglossini (Hymenoptera: Apidae). Journal of the Kansas Entomological Society 39(4):607-629. https://www.jstor.org/stable/25083566
Roberts, R.B. and Dodson, C.H. (1967). Nesting biology of two communal bees, Euglossa imperialis and Euglossa ignita (Hymenoptera: Apidae), including description of larvae. Annals of the Entomological Society of America 60(5):1007-1014. doi:10.1093/aesa/60.5.1007
xxxviiDressler, R.L. (1982). Biology of the orchid bees (Euglossini). Annual Review of Ecology and Systematics 13(1):373-394. doi:10.1146/annurev.es.13.110182.002105 https://www.researchgate.net/publication/234150443_Biology_of_the_Orchid_Bees_Euglossini
xxxviiiLogunov, D. (October 2020). Entomology Manchester. Orchid bees – flying jewels that pollinate flowers. https://entomologymanchester.wordpress.com/2020/10/15/orchid-bees-male-lives-matter/
xxxixDehon, M., Michez, D., Nel, A., Engel, M. S. and De Meulemeester, T. (2014). Wing shape of four new bee fossils (Hymenoptera: Anthophila) provides insights to bee evolution. PLoS ONE 9(10):e108865. doi:10.1371/journal.pone.0108865