

Alan Wade
The honey bee colony is almost always headed by a single queen irrespective of its natural or managed state. However finding a second queen is not uncommon. It routinely occurs when colonies are requeening themselves, that is when an old or deficient queen is being automatically replaced. Further there may be many queens in colonies preparing to reproduce in the process we call swarming.
The notion of running an extra laying queen to supercharge colonies has long lured apiarists into the belief that doing so would be straightforward. However maintaining a second queen has been so beset by the natural tendency of colonies to revert to the single-queen state that most beekeepers have abandoned all plans to do so.
Yet is any plan for running two-queen hives really so insurmountable, especially given we now know so much about their operationi? The quite recent development of the consolidated brood nest two-queen hive, where two queens are present in the same brood area, but kept apart by a queen excluder, has simplified their operation. If we add in the fact the normal swarming tendency of bees is strongly suppressed by the presence of a second queen – there are two lots of queen mandibular pheromone – one ponders why two-queen hives have not been in much wider use.
Nevertheless harnessing the power of a second laying queen is routinely practiced. Nucleus colonies kept to back up failing queens and splitting hives to prevent swarming are but two examples of running extra queens in separate colonies intended to maintain the strength of production hives. Such supernumerary colonies are either united at the commencement of the honey flow or as needed after disposing of a queen. This results in the combined forces of two laying queens being realised.
The history of two-queen history in outline
In the opening part of this series we examine the origin of the single queen hive, the exceptional circumstances under which a second queen may be maintained and efforts – dating back to the 1890s – to introduce and maintain a second queen. In practice, the operation of two-queen colonies has been chastened by their readiness to revert to the natural single queen condition.
Then, in Part II, we will visit the early pioneering efforts of some eminent bee researchers, notably Clayton Leon Farrar, Charles Harold Gilbert, Winston Edson Dunham, and John Edward Eckert and Frank Robert Shaw who developed techniques for establishing and operating two-queen hives. Their efforts, particularly in handling overly large colonies with a complex mixture of brood and honey supers, were heroic. While they were fully cognisant of the requirements to run such colonies, the skills to do so remained well beyond the capacity of most beekeepers.
In Part III we enter a period of successful operation of commercial apiaries operating with two-queen hives. We encounter the likes of Robert Banker (an editor of the beekeeping bible The Hive and the Honey Bee), New Zealand’s Department of Agriculture and Fisheries’ inventive G. M. Walton and Don Peer, a disciple of Farrar. They led the way in defining the limits of maintaining a second queen and in devising timely ways to harvest enormous honey crops. Until these beekeepers arrived, the large-scale operation of two-queen hives had been an elusive exercise, the territory of the most skilled apiarists.
Concluding Part IV will outline the development of the consolidated brood nest (CBN) hive. The CBN scheme replicates conventional single-queen hive management, brood below, honey supers on top. In so doing, Floyd Moeller and John Hogg revolutionised the operation of two-queen hives making them practical to operate. They introduced a range of innovative hive requeening techniques that are now helping address problems such as declining honey yields and increased disease prevalence. Hogg, in particular, revitalised the almost lost art of section comb honey production.
With this overview of two-queen hive history we can now turn to the genesis of single as well as two-queen honey bee colonies.
The antiquity of the honey bee colony headed by a single queen
While queens of all honey bees in the Apis genus mate with many drones, likely more than twenty for Apis melliferaii and even more for some other species, the monogamous (single-queen) condition is almost always adhered to. We start with an examination of the origins of the highly evolved eusocial honey bee colony, headed by a single queen, and will then review the rarer condition where a second queen is present.
Honey bees of the tribe Apini (Apis species) originated in the Miocene more than fifty million years ago. Their broader family (Apidae) includes the mainly eusocial corbiculate, so-called pollen basket, bees with a common progenitor dating back around eighty five million years (Figure 1).
Queens in both honey bees and other tribes of pollen basket bees evolved to co-opt the support of daughters to raise and provision brood. A small number of less related hallictid bees have independently developed a less advanced sociality where young larvae are mass provisioned in a common brood chamber.
These traits maximised the potential of the queen to successfully raise new generations of bees carrying her genes, in the more socially advanced species entirely removing the risks associated with her having to forage and provision her offspring. In a remarkable process of specialisation amongst bombinid and honey bees, the queen became the sole mother of all members of the colony. Stingless bees (Meliponini) can raise emergency queens and may have more than one laying queen – or queens — kept in reserve but they adopt the same principle of limiting the number of functional queensiii. The queen’s sterile female worker offspring, closely related to her, were relegated to the roles of foraging and of tending offspring although the workers themselves retained a capacity to exercise control over much of activity and behaviour of the colony. Such has been the degree of specialisation in these communities that subgroups, such as forager, swarm initiator and storage bees, may act independently of any queen signalling. Devolved decision making is one of the many hallmarks of advanced (eusocial) insects as well as in a few animal communities.
In the process of social specialisation both Apini (honey bee) and Meliponini (stingless bee) colonies became potentially immortal though, in practice wild and well as unmanaged colonies periodically fail. Only the individual bee is replaced making these insects a profoundly adaptable and efficient exploiter of floral resources. Further their propensity to accumulate stores has enabled them to radiate into more temperate climes and to survive extended periods of dearth.
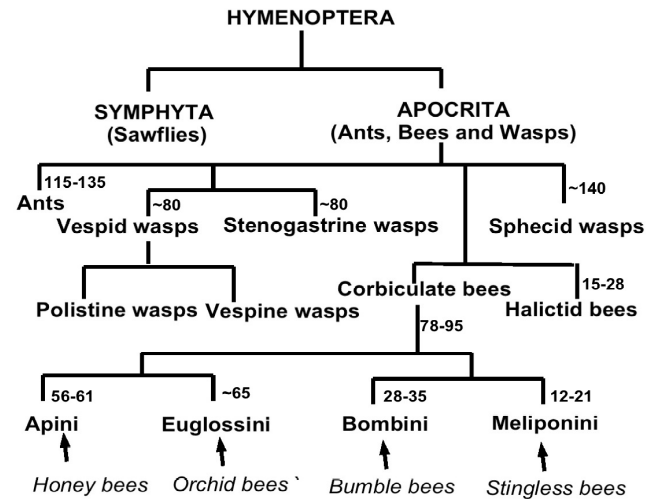
Figure 1 Simplified ancestral origin of social ants, bees and wasps (millions of years ago)
In all, eusociality has evolved at least eight times amongst the hymenoptera, four times amongst extant bee tribes: the advanced eusocial Apini (honey bees), the Meliponini (stingless bees), the primitive eusocial Bombini (bumble bees) and the now solitary or weakly social Euglossini (orchid bees) all dating back to a single ancestral primitively eusocial beeiv.
Honey and stingless bee tribes have evolved not only to a point where several generations of individuals coexist – that is where they work cooperatively to perpetuate the colony – but also to a condition where colony division (swarming) results in the formation of budded off colonies. Honey bees are more advanced in that they swarm to become immediately independent of the parent colony and in that the daughter colony of a primary swarm is headed by the parent colony queen. Within climatic limits and variability in availability of floral resources honey bees became capable of rapid and widespread dispersal, a situation limited amongst stingless bees where the daughter swarm only gradually becomes independent of the parent colony.
In the early ancestral condition, a single queen must have mated with a single drone, a strictly monogamous relationship. This condition was later relaxed so that the queen mated a number of times (polyandry)v, a trait particularly pronounced among honey bees (Table 1). At first the notion of a colony of ‘super sister’ worker bees working cooperatively with less related sisters to raise offspring, including future queens and drones only partly related to themselves, puzzled geneticists. However recognition that such colonies, comprising ‘super sister’ cohorts, would be more likely to survive prevailed. This was explained by the fact that selective pressures are on the honey bee colony – as expressed by the queen – not on the individual worker bee. The strategy of queens mating multiple times – to maximise sperm storage – and for each super sister cohort to express special attributes – for example better colony defence, superior nest hygienic behaviour, better comb building capability and better foraging facilityvi, – would better contribute to the survival of the colony and its queen.
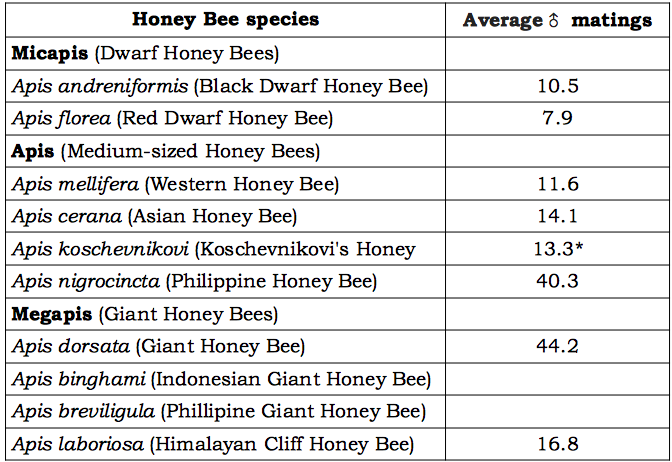
- More recent estimates suggest that thirteen drones may be needed to supply sufficient volume of sperm but that there may be many more matings.
Table 1 Honey bee polyandry
Wild colonies are small by comparison to those of contemporary commercial stock selected for honey gathering and pollination capability. For example, some African races of the Western Honey Bee (Apis mellifera) are poor comb builders and swarm prolifically, partly in response to pollen flowsvii. Indeed the reason that both the Western Honey Bee and the closely related Asian Honey Bee (Apis cerana) have been able to adapt to cold high latitude climates is that, in order to survive long winters, they both evolved to building their numbers quickly and to store surplus honey and pollen reserves. This aside, all honey bee colonies, for example the giant honey bees that migrate seasonally, must build quickly to reproduce in the process we call swarming.
Amongst the known eight or so extant honey bees, pheromonal signalling guides the maintenance of the single-queen colony, a feature likely characteristic of ancestral and now extinct species. The first record of the propensity of queens to fight and ensure that only a single queen remained was described in François Huber’s 1790 Letter IX to M. de Réaumurviii on the topic of the formation of swarms. Hugo von Buttell-Reepenix as well as Vivian Butz and Alfred Dietzx provide graphic and detailed accounts of the process of queen elimination and the circumstances under which it occurs. Hugo von Reepen famously reported:
If two queens come upon each other, only one will remain on the battlefield. If the queen is pleasing to every bee in every colony, the same thing may be said of the drones, who are extremely cosmopolitan, and who loaf about from hive to hive, and in consequence, apparently, of their specific odor, they are received peacefully everywhere, provided, of course, that the killing of drones has not yet begun…
Natural two-queen honey bee colonies
So we conclude that the normal condition for the honey bee is that of the colony being headed by a single queenxi. However queens, like workers and drones, all age and must be replaced periodically. So there are periods of changeover where many gynes (incipient queens) and drones (potential mates) are produced.
The droneright condition may persist all year but is generally restricted to times where queens can be raised or where nest conditions are particularly favourable. More typically colonies are perpetually queenright. However, and surprisingly, occasionally a multi-queen condition may be perpetuated. It is instructive to examine all the conditions under which queens are replaced to see how, and under what conditions, colonies maintain an additional queen.
The process of emergency queen replacement involves reprogramming a number of very young worker bees (typically up to around five less than one-day-old larvae) to become queen bees. This occurs with any sudden loss of the colony queen. Unlike queen cells started from scratch on the comb surface or on the comb margin in other more orderly processes of queen replacement, emergency queen cells show their tell-tale worker cell origin in the worker comb matrix. One characterising feature of emergency queen replacement is that there is always an extended period, a little over three weeks, during which the colony has no laying queen, that is where the colony is functionally queenless. Despite the setback the colony survives but only with a single replacement queen.
The process of supersedure, simple replacement of an ailing queen, involves the production of only 1-5 queen cells, sometimes but not always located near the centre of the brood nest. The surviving and newly mated daughter queen and mother, occasionally a total of three queensxii, may lay for an extended period, and is of fairly common occurrencexiii.
Now and then a second supersedure queen may result in a small swarm being cast with a virgin queen but this is an aberrant outcome. That both a founding and a daughter queen may coexistxiv is likely attributable to the fact that the old queen not only has low venom titre, and a low propensity to sting and hence defend herselfxv, but also a a produces weak pheromone signal, one that sometimes goes unrecognised by the young daughter queen. More often than not, however, the young queen prevails and the colony is returned to its single queen condition.
By 1921 reports of the presence of two queens in the one colony were so common that the editors of the British Bee Journal were prompted to exclaimxvi:
It is quite possible that there had been two queens in the hive, a condition that has been prevalent this year, or beekeepers are becoming more observant.
With supersedure, there is little or no interruption in the brood cycle and most often the old queen is lost either just before or just after the new queen is established. I have removed supersedure cells in colonies a number of times only to have them rebuild a few more cells on each occasion. For example Doolittlexiv, records:
After I had this experience with the colony that had ‘two Queens in a hive’, (which was a surprise to so many fifteen years ago, when it was thought that no colony ever tolerated but one laying queen at a time), I began to watch for a like circumstance to occur, which happened about a year from that time. In the latter case, as soon as I found the cells, they were sealed over, and not knowing just when they would hatch [emerge], I at once cut them out and gave them to nuclei. In a few days I looked in the hive again, when I found more cells started, which were again cut off and given to nuclei, just before it was time for them to hatch [emerge]. In this way I kept the bees from their desired object for some two months, or until I saw that the old Queen was not going to live much longer, when I left one of the cells, which they had under headway, to mature. By this plan I got about sixty as fine Queens as I ever reared, and laid the foundation for my present plan of securing Queens, which is about to be given in this book.
In several instances I found that the old queen continued to lay well and an emerged daughter queen failed to replace her mother possibly signalled by a vastly improved late spring honey flow condition. Hepburn and Radloffxvii record instances where the two-queen status of colonies persists in African races of the Western Honey Bee including the Cape Honey Bee Apis mellifera capensis, the Arabian Honey Bee Apis mellifera jemenitica and in the African Honey Bee Apis mellifera scutellataxviii. Mathisxix records uncharacteristic supersedure two-queen-colony persistence in the Tellian or Tunisian bee – also called Punics (Apis mellifera intermissa).
The process of swarming results in the production of a large number of potential queensxx, though the periodic formation of queen cell cups resulting from crowding is unrelated and arises from buildup of footprint pheromonexxi. Apis mellifera lamarckii (Lamarck’s Honey Bee) is famous for producing between several dozen and several hundred queen cellsxxii. Depending on the number of swarms issued, the parent colony will also experience a brief period where there is no laying queen or indeed may swarm itself to death. Nevertheless is not uncommon for there to be number of nearly fully developed pre-emergent queens and even a number of virgin queens present once the first (prime) swarm departs.
Surfeit virgin queens remaining in the parent colony after all swarming has ceased are quickly eliminated so that, in line with the evolved condition, only one queen survives. After swarms issuing with a number of virgin queens may split and settle as independent new colonies again each establishing as a single queen unit. The overall swarming process is described cogently by Demuthxxiii.
Apart from the normally temporary arrangement of two-queen colonies arising from queen supersedure, there are appear to be no scenarios amongst European honey bees where two-queen colonies arise spontaneously as a result of queen replacement. In a remarkable insight into the differences between swarming and supersedure Hogg notedxxiv:
Queen supersedure is never the cause of swarming. But supersedure may occur concurrently with the swarm whenever the queen’s failure was caused by being over-taxed while generating the bees for that swarm. Significantly, it is the virgin queen that is then selected to accompany the swarm. The failed queen is retained in the parent hive to be superseded in turn…
Apparently, just as the survival instinct of the bees in control won’t allow a swarm to leave a hive without a replacement queen in the parent [hive], they won’t allow a failing queen to issue with that swarm.
While the propensity of any colony to default to a single queen state is always very strong, it was such insight that eventually allowed John Hogg and, earlier, Eugene Killionxxv to devise methods of crowding bees so that that were unlikely to swarm. As we shall see in the concluding part to this series, Hogg was able to devise a scheme of running stable two-queen colonies where the natural antagonism between queens was eliminated and where the cooperative foraging behaviour amongst worker bees was optimised.
Parthenogenic requeening
In rare instances workers will spontaneously produce fertile offspring in a process termed thelytokousxxvi parthenogensisxxvii. Such queenless colonies may and often do requeen themselves. Even in this circumstance only a single-queen survives.
Mackensonxxviii and Butlerxxix document a record of such instances with the Italian Honey Bee, Apis mellifera ligustica, the Caucasian Honey Bee Apis mellifera caucasia, the Syrian Honey Bee, Apis mellifera syriaca, the Tunisian Honey Bee, Apis mellifera intermissa and the Cape Honey Bee, Apis mellifera capensis from South Africa. The trait is so pronounced in the Cape Honey Bee that, according to Butler, colonies that become queenless are reluctant to raise emergency queens. Such colonies continue to function normally, laying workers – pseudo queens – producing both normal workers and small drones raised in regular worker comb. However under normal circumstances, greater than 99% of European races of the Western Honey Bee colonies will dwindle and perish as will many Cape Honey Bee colonies. Interestingly drones from hopelessly queenless colonies in all honey bee colonies retain the potential to pass on the former queen’s genes, that is assuming they can then mate successfully.
Smith (1961)xxx makes the earliest reference to the existence of fertile laying workers in the Tellian Honey Bee (Apis mellifera intermissa):
Towards the end of this period, bees from Tunis, which were referred to as Punics, caused some interest in Britain, where they were first imported by J. Hewitt, who wrote under the name ‘A Hallamshire Beekeeper’. John Hewitt (1892) reported that Punic bees could rear queens from the eggs of laying workers, and this appeared to be quite a common occurrence with his strains.
Apart from many confused reports disputing the existence, temperament and productivity of these ‘Punic bees’ (Apis mellifera intermissa) outside their native Tunisia and in the near Middle East, Hewittxxxi made findings of these parthenogenic bees almost a decade earlier though Hewitt’s claims were later roundly dismissedxxxii. This was not a case of workers laying and having queens raised parthenogenetically but rather workers laying multiple eggs in a cell alongside fertile queens from which drones are raised in worker cells. It appears that this observation was wrongly ascribed the queen to being a drone layer.
The more celebrated discovery of parthenogenesis amongst Cape Honey Bees (Apis mellifera capensis) dates mainly from the detailed observations of Onionsxxxiii. His findings, made between 1912 and 1914, attracted wide commentaryxxxiv, but it was not until the 1950s that any systematic reviewsxxxv of Cape Honey Bee biology were undertaken. The well-respected Baldenspergerxxxvi gave no credence to the Punic or Tunisian Honey Bee (Apis mellifera intermissa) or the Syrian Honey Bee (Apis mellifera syricia) being able to requeen parthenogenetically.
It is very clear from the 1857 writings of the eminent physiologist Carl Theodor Ernst von Sieboldxxxvii known well to Johann Dzierżońxxxviii, the ‘Father of Beekeeping’, that such fertile unmated queens were entirely unknown to the beekeeping literatii, that is prior to the discovery of the aberrant self-queening behaviour in the Cape Honey Bee.
Recombinant primary swarms, those merging from different colonies, each containing an old queen occasionally settle to form a new colony headed by two laying queens. This condition appears rare and such swarm colonies will eventually revert to the single queen colony statusxxxix.
Parasitising two-queen colonies. The existence of two-queen colonies and the aforementioned self-requeening of queenless colonies is pronounced in some races of African strains of the Western Honey Bee, particularly in the aforementioned Cape Honey Bee, Apis mellifera capensisxl. Single queen Cape Honey Bee colonies (or queens) also drift to or swarm into African Honey Bee (Apis mellifera scutellata) colonies sometimes resulting in the formation of two-queen coloniesxli. There the African Honey Bee queen is relegated to the nest periphery and the colony is overtaken by Cape Honey Bee laying workers. Such colonies, and indeed whole African Honey Bee apiaries, have perishedxlii leading to controls being placed on Cape Honey Bee migratory beekeeping.
Magnumxliii provides some limited evidence that late swarms of European honey bees supplant existing honey bee colonies or progressively rob them thus parasitising their resources. However this kleptoparasitism does not lead to the formation of two-queen-colonies. This phenomenon has been most widely reported on for takeover of the West African Honey Bee (Apis mellifera scutellata) by the Cape Honey Beexliv.
The reproductive biology of these southern African races of honey bee is complex but the fact that two-queen colonies commonly exist confirms that multiple-queen colonies may, a priori, be operated successfully even where queen excluders are not used.
Amongst normal managed colonies employing queen excluders, the appearance of a second, or even a third laying queen, is common where brood nests are split and where a second hive entrance is provided allowing an additional queen to be raised and to mate. This sometimes occurs in practicing Demaree swarm control where brood, separated from the queen, raises queens one of which may mate and establish a separate brood nest.
Early production hives containing two queens
From 1892, Wells and othersxlv had operated back-to-back single-queen hives using their combined work forces to boost honey production. They positioned honey supers above a queen excluder spanning both brood nests, a setup of independent single queen hives sharing common storage space.
In a historic move in 1907, Alexanderxlvi had pioneered operation of single hives containing two or more queens. He employed an elaborate engorgement and preconditioning technique to introduce an extra queen to an already queenright colonyxlvii, a technique that Hallxlviii was soon to experiment with. Others, such as Hand, Sherrod, Chambers, Wright, Bussy, Gray and Joice were soon to followxlix.
In a footnote to a 1911 letter by A.B. Marchant deprecating the two-queen process, the Gleanings in Bee Culture editorsl attributed much of the success of Alexander’s and Doolittle’s pursuits – including those of operating hives with additional queens – to their consummate beekeeping skills and to the extraordinary floral resources of their beekeeping localities. Alexander is credited with harvesting over thirty seven tonnes of honey from 750 hives located at a single site so could rail against the prevailing belief that multi-queen colonies could not be maintained.
Also around 1907, and in a concurrent pushli, Cruadh, Ellis and Medicus were to establish a more successful approach to two-queen hive beekeeping in the United Kingdom. By employing queen excluders – at least one – a succession of beekeepers, Hall (1907), Cruadh (1907), Ellis (1908), and Medicus (1910) were all able to establish a second queen and reliably maintain that condition through to the end of the main honey flow. Cruadh provided the most explicit details, clearly delineating the doubled hive system – separate hives sharing common honey supers – from the two-queen system – two queens in the one colony, both illustrating and describing in detail the set up and operation of two-queen hives. His lucidly documented account – spread over a number of 1910 issues of the British Bee Journal – makes him, if not the original inventor, the stand out early successful two-queen hive operator.

Figure 2 Marchant’s apiary on the Appalachicola River in Floridalii
But these were early forays and it was not until the 1930s that Farrar, and later Moeller and Hogg, developed more ingenious methods for running powerful two-queen hives, robust schemes with very general application. The American research effort, supported by the United States Department of Agriculture, was much more sustained than that of the southern Scottish venture and found universal application.
The take home message from all this early experimentation was that two-queen colonies may persist in that state, even after removal of all queen excluders, that was at least until autumn or until the main honey flow was over. Spojaliii in Yugoslavia, corresponding with Kovtunliv in the Kharkov area of the then USSR independently explored the conditions under which multiply queened hives could be established and maintained in the absence of queen excluders. Their findings, referenced by Farrarlv confirmed the Alexander’s notion of hives hives operating with ‘a plurality-of-queens’.
By the 1920s it became apparent that the conditions of finding two queens in the one honey bee colony were contingent upon either:
- colonies undergoing queen supersedure where the hive tolerated mother and daughter sometimes, but not always, for an extended period of time; or
- hive manipulation involving separation of brood leading to a second laying queen. Both daughter, or occasionally two daughters, may each lay in discrete brood nests for a period especially if the daughter unit is provided a separate flight entrance and the nests are separated by a queen excluder; or
- a more determined effort to establish a second queen – usually a laying queen – as demonstrated by Alexander and Kovtun – where her introduction and establishment were carefully finessed.
While the latter technique was to await full development by Farrar in the 1930s, a spate of records of heather hunting beekeepers, notably Medicus and Ellis, noted above, were to develop and operate powerful two-queen colonies between 1910 and the 1920s.
The full conditions for queen acceptance to any honey bee colony were fully investigated and articulated by Hogg. His basic finding was that the condition of acceptance of two or more queens were essentially the same as those for acceptance of one queenlvi. A colony made gyne free – that is without any potential queen – will, depending on the setup and prevailing conditions, readily accept two or more queens. The question then became one of maintaining any additional queen or queens.
It soon became apparent that, particularly under honey flow conditions, queens and her workers may be too preoccupied laying to supply a honey gathering workforce, to be actively patrolling their larger colony. Such queen introductions, practiced in the lead up to a honey flow, increased honey production by simple dint of even the temporary presence of a second laying queen.
However large colonies run on good honey flows reliably maintain two queens. At the end of the season, and without intervention, each colony will almost invariably revert to the natural single-queen condition. This often reported finding was confirmed in my experience of running two-queen colonies over the very poor seasons of 2018-2019 and 2019-2020. Of nine colonies, five reverted to single-queen hives during repeated stop-start honey flows. While a second queen was successfully restored in four of those colonies, the final queen loss occurred too late in the season to justify requeening but only one of the nine colonies performed well. Clearly two-queen hives are unstable in any period of dearth, not just at end of the season. A great deal still remains to be understood about the stability and operation of two-queen colonies.
Despite the overwhelming evidence that two-queen hives are far more productive than equivalent pairs of single-queen hives, in a poor season this is not always the case. Lensky and Golanlvii report that in drought years only marginally more honey is produced than in a single-queen colony. Importantly they also report that:
…each one of the two queens in a two-queen colony oviposited less than one queen in a single-queen hive.
Artificial multiple-queen colonies
More recently multiple queen colonies, those operated without a queen excluder, have been operated year round albeit after disabling queen defences. For example, Paleolog and coworkerslviii clipped queen stings and Zheng and fellow collaboratorslix ablated, that is blunted, the mandibles of queens. Using these queens, Zheng’s group were able to maintain 3-6 queen colonies over winter. Of more interest has been the finding that multiple queens can be maintained in colonies by masking chemicals daubed onto mandibles and antennaelx.
The general condition of multi-queen colonies have been investigated by Lensky and Darchenlxi, though the idea of clipping queen stings to maintain a second queen goes well back to the aftermath of Alexander’s findings where masking techniques – e.g. smoking bees – were employed to introduce additional queenslxii.
Woykelxiii and by Szabo, Wyborn and many otherslxiv have studied the successes and problems of queen banking, notionally multiple queen colonies, including keeping mated queens over winter. However, queens under these conditions are always confined and, while in a potential laying condition, are functionally unable to do so. Other studies such as those conducted by Dietzlxv have focussed on problems of year round maintenance of two-queen colonies.
Expansive studies on the mating behaviour of queens and of queens, both within and introduced to hives, made by Garylxvi and Szabolxvii provide some clue to the behaviours of colonies with more than one queen and signal some of the problems attendant to maintenance of two-queen hives. For example Farrarlxviii notes:
Swarming is less of a problem in two-queen colonies than in strong single-queen colonies, but queen cells started because of a failing queen or crowding in either brood nest will stimulate production of queen cells in the other.
In Part II of this series, we will trace the major developments in operating two-queen honey bee colonies commencing with the findings of Farrar. Despite all schemes employing two queens demonstrating greatly enhanced honey production, every scheme presented logistic challenges that have only been overcome relatively recently. The truly modern and efficient two-queen hive is the topic that will be covered in concluding Part IV.
Readings
iWade, A. (2019a). Establishing two queens instead of one queen in a honey bee colony Part I. Principles of introducing and running two-queen colonies. The Australasian Beekeeper 120(8):18-21.
Wade, A. (2019b). Establishing two queens instead of one queen in a honey bee colony Part II. Setting up two-queen colonies. The Australasian Beekeeper 120(9):16-21.
Wade, A. (2019c). Establishing two queens instead of one queen in a honey bee colony Part III. Operating two-queen colonies. The Australasian Beekeeper 120(10):22-25.
iiTarpy, D.R., Delaney, D.A. and Seeley, T. (2015). Mating frequencies of honey bee queens (Apis mellifera L.) in a population of feral colonies in the northeastern United States. PLoS ONE 10(3):e0118734 https://doi.org/10.1371/journal.pone.0118734
iiiBradbear, N. (2009). Bees and their role in forest livelihoods: A guide to the services provided by bees, Chapter 6. Meliponiculture of stingless bees. FAO Rome. http://www.fao.org/3/i0842e/i0842e00.htm
ivCardinal, S. and Danforth, B.N. (2011). The antiquity and evolutionary history of social behavior in bees. PLoS ONE 6(6): e21086. https://doi.org/10.1371/journal.pone.0021086
vPalmer, K.A. and Oldroyd, B.P. (2000). Evolution of multiple mating in the genus Apis. Apidologie 31:235–248. https://www.apidologie.org/articles/apido/pdf/2000/02/m0203.pdf
viNowak, M.A., Tarnita, C.E. and Wilson, E.O. (2010). The evolution of eusociality. Nature 466(7310):1057–1062. https://doi.org/10.1038/nature09205 and https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3279739/
viiHepburn, H.R. and Radloff, S.E. (1998). Honeybees of Africa. Springer-Verlag. Berlin, New York, 370 pp., p.139.
viiiHuber, F. (1806). New observations on the natural history of bees, Volume I by François Huber, Letters to M. Bonnet. [Letters written between 1787 and 1791.] http://www.bushfarms.com/huber.htm
ixvon Buttel-Reepen, H. (1900). Sind die bienen reflexmaschinen? Experiementelle beiträge zur biologie der honigbiene. Biologisches Centralblatt 20(1):1-82. Are bees reflex machines? Experimental contribution to the natural history of the honey-bee translated by Mary H. Geisler. Medina, The A.I. Root Co., 1907. 51pp., p.10. https://babel.hathitrust.org/cgi/pt?id=hvd.hn4t52;view=1up;seq=19
xButz, V.M. and Dietz, A. (1994). The mechanism of queen elimination in two-queen honey bee (Apis mellifera L.) colonies. [Nouvelles Observations Sur Les Abeilles]. Journal of Apicultural Research 33(2):87-94. https://doi.org/10.1080/00218839.1994.11100855
Dietz, A. (1986). Theories on two-queen colonies. American Bee Journal 126(2):84, 133. Cited by Butz and Dietz (1994). loc. cit.
Jones, C.L. (1959). A two-queen colony that pays. Gleanings in Bee Culture 87(5):276-277. Cited by Butz and Dietz (1994). loc. cit.
xiWilson, E.O. and Hölldobler, B. (2005). Eusociality: Origin and consequences. Proceedings of the National Academy of Sciences of the United States of America 102(38):3367–3371. https://www.pnas.org/content/102/38/13367
xiiFarrar, C.L. (1953). Two-queen colony management. American Bee Journal 93(3):108-110, 117 reprinted as Farrar, C.L. (1953). Two-queen colony management. Bee World 34(10):189-194. https://doi.org/10.1080/0005772X.1953.11094821
xiiiSee Doolittle, G.M. (1889). Scientific queen-rearing as practically applied being a method by which the best of queen-bees are reared in perfect accord with nature’s ways, 184pp., p.27. Chicago, Ills. Thomas G. Newman & Son, 923 & 925 West Madison Street. https://ia800200.us.archive.org/12/items/cu31924003263724/cu31924003263724.pdf
von Buttel-Reepen (1900), p.10. and
Spoja (1953), p.195 loc. cit. and
Phillips, E.F. (1915). Beekeeping: A discussion of the life of the honeybee and of the production of honey. The Rural Life Sciences, Ed. Bailey, L.H. https://victoriancollections.net.au/media/collectors/51d110e42162ef12e06aa06b/items/5341f5212162ef0a845d5a79/item-media/5341f6e42162ef0a845d648d/original.pdf
xivButler, C.G. (1954a). The World of the Honeybee, p.60 and plate 18, p.99. Collins New Naturalist, London. Collins, St James Place, London.
Butler, C.G. (1954b). The method and importance of the recognition by a colony of honeybees (A. mellifera) of the presence of its queen. Transactions of the Royal Entomological Society of London 105(pt. 2):11-29. https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1365-2311.1954.tb00773.x
Butler, C.G. (1957). The process of queen supersedure in colonies of honeybees (Apis mellifera L.). Insectes Sociaux 4(3): 211-223. https://page-one.springer.com/pdf/preview/10.1007/BF02222154
Butler, C.G. (1960). The significance of queen substance in swarming and supersedure in honey-bee (Apis mellifera L.) colonies. Proceedings of the Royal Entomological Society of London. Series A, General Entomology 35(7‐9):129-132. https://doi.org/10.1111/j.1365-3032.1960.tb00681.x Reprinted in April 2009 at Physiological Entomology 35(7-9):129–132.
Neumann, P., Pirk, C.W.W., Hepburn, H.R. and Radloff, S.E. (2001). A scientific note on the natural merger of two honey bee colonies (Apis mellifera capensis). Apidologie 32(1):113-114. https://doi.org/10.1051/apido:2001116
Simpson, J. (1958). The factors which cause colonies of Apis mellifera to swarm. Insectes Sociaux 5(1):77-95. https://doi.org/10.1007/BF02222430 Simpson, J. (1958). The problem of swarming in beekeeping practice. Bee World 39(8):193-202. https://doi.org/10.1080/0005772X.1958.11095063 and https://www.tandfonline.com/doi/abs/10.1080/0005772X.1958.11095063
xvCollison, C. (2018). Venom/venom glands – A closer look. The Australasian Beekeeper 120(3):46-48.
xviBrichter, G. (1921). Two queens in a hive. British Bee Journal and Beekeeper’s Adviser 49:384. https://ia800203.us.archive.org/17/items/britishbeejourna1921lond/britishbeejourna1921lond.pdf
xviiHepburn and Radloff (1998). Chapter 5, p.140, loc. cit.
xviiiClaus, B. (1984). Ex Africa: Botswana, some observations on biology and behaviour. South African Bee Journal 56(5):113-116. Cited by Hepburn and Radloff (1998), p.140. loc. cit.
xixMathis, M. (1952). Polygyny, temporary but constant and natural, in Apis mellifica var. punica in Tunisia; Absence of mortal combat among the queens. Comptes rendus hebdomadaires des séances de l’Académie des sciences 234(24):2390-2392. PMID: 12979358 [Note Mathis’s reference is probably to a variant of the local Tellian Honey Bee, Apis mellifera intermissa.] https://www.ncbi.nlm.nih.gov/pubmed/12979358
Mathis, M. (1952). Temporary (but natural and maintained) polygyny in A. mellifera var. punica in Tunisia: Queens not killing each other. Gazette Apicole 53(548):230-23. Apicultural abstract 134/53. Cited by Walton (1974). loc.cit.
xxWinston, M.L. (1980). Swarming, after swarming, and reproductive rate of unmanaged honeybee colonies (Apis mellifera). Insectes Sociaux 27(4):391-398. https://link.springer.com/article/10.1007/BF02223731 Winston, M.L., Taylor, O.R. and Otis, G.W. (1980). Swarming, colony growth patterns, and bee management. American Bee Journal 120(12):826-830.
xxiLensky, Y. and Slabezki, Y. (1981). The inhibiting effect of the queen bee (Apis mellifera L.) foot-print pheromone on the construction of swarming queen cups. Journal of Insect Physiology 27(5):313-323. https://doi.org/10.1016/0022-1910(81)90077-9
xxiiHepburn, H.R. and Radloff, S.E. (1998).p.139. loc. cit.
xxiiiDemuth, G.S. (1921). Swarm control. Farmers Bulletin 1198:1-45. https://ia601401.us.archive.org/32/items/CAT87202908/farmbul1198.pdf
Demuth, G.S. (1917). Commercial comb-honey production. Farmers Bulletin 1039, United States Department of Agriculture, 40pp. https://ia902309.us.archive.org/19/items/CAT87202962/farmbul1039.pdf
xxivHogg, J.A. (2006). Colony level honey bee production: The anatomy of reproductive swarming. American Bee Journal 146(2):131-135. http://www.twilightmd.com/Samples/Hogg/Hogg_Halfcomb___Publications/ABJ_2006_February.pdf
xxvKillion, E.E. (1981). Honey in the Comb. Dadant & Sons. Inc, Carthage, Illinois.
xxviEsko Suomalainan describes three types of parthenogenesis: Arrhenotoky where unfertilised eggs develop pathenogenetically into males; Thelytoky where unfertilised eggs develop into females; and Deuterotoky where unfertilsed eggs develop into both sexes. Suomalainen, E. (1950). Parthenogenesis in animals. Advances in Genetics3:193-253. https://doi.org/10.1016/S0065-2660(08)60086-3
xxviiGoudie, F. and Oldroyd, B.P. (2014). Thelytoky in the honey bee. Apidologie 45(3):306–326. https://link.springer.com/article/10.1007/s13592-013-0261-2
Oldroyd, B.P., Allsopp, M.H., Gloag, R.S., Lim, J., Lyndon A. Jordan, L.A. and Beekman, M. (2008). Thelytokous parthenogenesis in unmated queen honeybees (Apis mellifera capensis): Central fusion and high recombination rates. Genetics 180(1):359-366. https://doi.org/10.1534/genetics.108.090415 and https://www.genetics.org/content/180/1/359.full
Beekman M., Allsopp M.H., Lim J., Goudie F. and Oldroyd, B.P. Asexually produced Cape honeybee queens (Apis mellifera capensis) reproduce sexually, Journal of Heredity 102(5):562-566. https://10.1093/jhered/esr075 Full text at https://www.researchgate.net/publication/51508237_Asexually_Produced_Cape_Honeybee_Queens_Apis_mellifera_capensis_Reproduce_Sexually
Pirk, C., Lattorff, H., Moritz, R., Sole, C., Radloff, S., Neumann, P., Hepburn, H., Crewe, R., Beekman, M., Allsopp, M., Lim, J., Goudie, F. and Oldroyd, B. (2012). Reproductive biology of the Cape Honeybee: A critique of Beekman et al. Journal of Heredity 103(4):612-614. https;//doi.org/10.1093/jhered/ess007
Beekman, M., Allsopp, M.H., Lim, J., Gouldie, F. and Oldroyd, B. (2012). Response to ‘Reproductive biology of the Cape Honey Bee: A critique by Beekman et al.’ by Pirk et al. Journal of Heredity 103(4):614-615.
xxviiiMackenson, O. (1943). The occurrence of parthenogenic females in some strains of honey bees. Journal of Economic Entomology 36(3):465-467. https://doi.org/10.1093/jee/36.3.465
xxixButler, C.G. (1954a). loc. cit. pp.57-58.
xxxSmith, F.G. (1961). The races of honeybees in Africa. Bee World 42(10):255-260. https://doi.org/10.1080/0005772X.1961.11096896
xxxiHewitt, J. (1883). Syrian queens and fertile workers. British Bee Journal and Bee-keepers’ Record and Adviser 11(124):66-67. https://biodiversitylibrary.org/page/25729174
Hewitt, J. (1884). New races of bees – fertile workers. American Bee Journal 20(19):294-295. https://ia802702.us.archive.org/22/items/americanbeejourn2084hami/americanbeejourn2084hami.pdf
A Hallamshire Beekeeper [pseudonym for John Hewitt] (1891). Qualities of the African, or Punic, bee. American Bee Journal 28(22):701-702. https://www.biodiversitylibrary.org/item/77776#page/515/mode/1up Cited and refuted in detail by Baldensperger (1918), loc. cit. pp.375-376.
Hewitt, J. (1892). Fertile workers—their utility. Journal of Horticulture 25(3):139. Cited by Ruttner and Mackensen (1952). loc. cit. and by Rinderer, T.E. (1986). Bee Genetics and Breeding. USDA Baton Rouge. Academic Press Inc. Harcourt Brace Jovanovich, Publishers. p.229. https://books.google.com.au/books?id=MyTgBAAAQBAJ&pg=PA229&dq=Hewitt,+J.+(1892).+Fertile+workers%E2%80%94their+utility.&hl=en&sa=X&ved=0ahUKEwi2_dGd6pjmAhWBX30KHQLdCtIQ6AEIKTAA#v=onepage&q=Hewitt&f=false
xxxiiCowan, T.W. (1891). Topics of interest: English editors and punic bees. American Bee Journal 28(24):747-749.
Cowan, T. (1891). Punic and Minorcan bees – explanation. American Bee Journal 28(26):811-813. https://ia902609.us.archive.org/23/items/americanbeejourn28hami/americanbeejourn28hami.pdf
xxxiiiOnions, G.W. (1912). South African fertile-worker bees. Agricultural Journal of the Union of South Africa 3(5):720-728. https://journals.co.za/content/ajusa/3/5/AJA0000021_1356?fromSearch=true
Onions, G.W. (1914). South African fertile-worker bees. Agricultural Journal of the Union of South Africa 7:44-46. https://journals.co.za/content/ajusa/7/1/AJA0000021_317?fromSearch=true
Jack, R.W. (1917). XXI. Parthenogenesis amongst workers of the Cape Honey Bee: Mr G.W. Onions’ experiments. Transactions of the Royal Entomological Society of London 64(3-4):396-403. https://www.biodiversitylibrary.org/item/55138#page/14/mode/1up
xxxivvan Warmelo, D.S. (1912). South African fertile-worker bees and parthenogenesis. Agricultural Journal of the Union of South Africa 3:786-789. https://journals.co.za/content/ajusa/3/6/AJA0000021_611?fromSearch=true
Nachtsheim, H. (1912). Parthenogenese, eireifung, und geschlechtsbestimmung bei der honigbiene. Sitzungsber Ges Morph Physiol München 28:22-29. Cited by Nachtsheim (1913). loc. cit.
Nachtsheim, H. (1913). Cytologische studien über die geschlechtsbestimmung bei der honigbiene (Apis mellifica L.). Archiv für Zellforschung 11:573-608. http://honeybee.drawwing.org/node/971 Cited by Oldroyd et al (2008). loc. cit.
Anderson, J. (1918). Laying workers which produce female offspring. American Bee Journal 58(6):192. https://ia800907.us.archive.org/21/items/americanbeejourn5859hami/americanbeejourn5859hami.pdf
Gough, L.H (1928). Apistischer brief aus Südafrika. Der Bienenvater 60:30-32. Account of the biology of the honeybee in the Eastern Cape covering the essential features of capensis: A queenless swarm can produce a new queen. Cited by Hepburn, S.G. and Guye, H.R. (1993). An annotated bibliography of the Cape Honeybee, Apis mellilera capensis Eschscholtz (Hymenoptera: Apidae). African Entomology 1(2):235-252. https://journals.co.za/content/ento/1/2/AJA10213589_64?fromSearch=true and https://journals.co.za/docserver/fulltext/ento/1/2/65.pdf?expires=1575160431&id=id&accname=guest&checksum=141EB0D11CFFEF15A756D3F4C48BDB75
xxxvFyg, W. and Hodges, F.E.D. (1950). Can workers and queens of the honeybee be raised from unfertilised eggs? Bee World 31:17-19. https://www.tandfonline.com/doi/abs/10.1080/0005772X.1950.11094622?journalCode=tbee20
Ruttner, F. and Mackensen, O. (1952). The genetics of the honeybee. Bee World 33(5):671-679. https://www.tandfonline.com/doi/abs/10.1080/0005772X.1952.11094729 Tucker, J.W. (1958). Automictic parthenogenesis in the honey bee. Genetics 43(3):299-316. www.genetics.org/content/genetics/43/3/299.full.pdf
xxxviBaldensperger, P.J. (1918). Punics and parthenogenesis. American Bee Journal 58(11):375-376. https://ia800907.us.archive.org/21/items/americanbeejourn5859hami/americanbeejourn5859hami.pdf
xxxviivon Siebold, C.T.T. (1857). A true parthenogenesis in moths and bees: A contribution to the history of reproduction in animals,125pp. Translated by William S. Dallas, F.L.S. &c. London: John van Voorst, Paternoster Row. True parthenogenesis in the honey-bee pp.38-91. https://ia802302.us.archive.org/22/items/ontrueparthenoge00sieb_0/ontrueparthenoge00sieb_0.pdf
For a broader overview of parthenogenesis see Uyenoyama, M.K. (1984). On the evolution of parthenogenesis: A genetic representation of the cost of meiosis. Evolution 38(1):87-102. https://doi.org/10.1111/j.1558-5646.1984.tb00262.x
xxxviiiDzierżoń (1882). Dzierzon’s rational beekeeping or The theory and the practice of Dr Dzierzon of Carlsmarkt. (Translated by H. Dieck and S. Stutterd; Edited and reviewed by Charles Nash Abbott. Houlston & Sons, Paternoster Square, Southall: Abbott Bros. from Jan Dzierzon’s (1852) treatise Nachtrag zur Theorie und Praxis des neuen Bienenfreundes; oder, Einer neuen Art der Bienenzeucht mit dem günstigsten Erfolge angewendet und dargestellt von Dzierzon. Nördlingen, Beck.0 https://ia800201.us.archive.org/17/items/dzierzonsration00stutgoog/dzierzonsration00stutgoog.pdf
xxxixReported by former Canberra Region Beekeepers club president Cormac Farrell in the summer of 2015-2016.
xlHepburn and Radloff (1998), pp.140-141. loc. cit.
xliHepburn, H.R. (2001). The enigmatic Cape Honey Bee. Bee World 82(4):181-191. https://www.tandfonline.com/doi/abs/10.1080/0005772X.2001.11099525
Muerrle, T.M. (January 2008), PhD Thesis. Queens, pseudoqueens and laying workers: Reproductive competition in the Cape Honeybee (Apis mellifera capensis Eschscholtz). https://core.ac.uk/download/pdf/11984223.pdf
xliiClauss, B. (1983). Bees and beekeeping in Botswana, Ministry of Agriculture, Gaborone. Cited in Hepburn and Guye (1993). loc. cit.
xliiiMagnum, W.A. (1 May 2009). Usurpation: A colony taken over by a foreign swarm. American Bee Journal 149(5). https://americanbeejournal.com/usurpation-a-colony-taken-over-by-a-foreign-swarm/
Magnum, W.A. (2010). The usurpation (takeover) of established colonies by summer swarms in Virginia. American Bee Journal 150(12):1139-1144. Cited by Oliver, R. (2014). What’s happening to the bees? – Part 2. http://scientificbeekeeping.com/whats-happening-to-the-bees-part-2/
Magnum, W.A. (2012). Colony takeover (usurpation) by summer swarms: They chose poorly. American Bee Journal 153(1):73-75.
Magnum, W.A. (1 January 2018). Usurpation: a colony taken over by a foreign swarm. American Bee Journal 158(1). https://americanbeejournal.com/usurpation-colony-take-summer-swarms/
xlivSchneider, S. Deeby, T., Gilley, D. and DeGrandi-Hoffman, G. (2004). Seasonal nest usurpation of European colonies by African swarms in Arizona, USA. Insectes Sociaux, 2004 51(4):359-364. https://doi.org/10.1007/s00040-004-0753-1
Danka, R.G., Hellmich, R.L. and Rinderer, T.E. (1992). Nest usurpation, supersedure and colony failure contribute to Africanization of commercially managed European honey bees in Venezuela. Journal of Apicultural Research 31(3-4):119-123. https://doi.org/10.1080/00218839.1992.11101272
xlvBritish Bee-Keepers’ Association. Quarterly Converzation (April, 7, 1892). British Bee Journal, Bee-Keepers’ Record and Adviser 20:132-133. https://ia802508.us.archive.org/29/items/britishbeejourna1892lond/britishbeejourna1892lond.pdf
Wade, A. (2019d). Doubling hives: Part I Doubled vs two-queen hives. Canberra Region Beekeepers Newsletter https://actbeekeepers.asn.au/bee-buzz-box-december-2019-doubling-hives-.part-i-doubled-vs-two-queen-hives/
Wade, A. (2020a). Doubling Hives: Part II The Wells System. Canberra Region Beekeepers Newsletter https://actbeekeepers.asn.au/bee-buzz-box-january-2020-doubling-hives-.part-ii-the-wells-system/
xlviAlexander, E.W. (1907a). A plurality of queens without perforated zinc: How the queens are introduced: The advantages of the plural-queen system. Gleanings in Bee Culture 35(23):1136-1138. https://babel.hathitrust.org/cgi/pt?id=umn.31951d00953180r;view=1up;seq=660
Alexander, E.W. (1907b). The plural queen system: All queens but one disappear at the end of the season. Gleanings in Bee Culture 35(17):1496. https://babel.hathitrust.org/cgi/pt?id=hvd.32044106180268;view=1up;seq=562
Alexander, E.W. (1908). The plural two queen system: Is it profitable for bee-keepers to practice it? Gleanings in Bee Culture 36(18):1135. https://babel.hathitrust.org/cgi/pt?id=uc1.b3458203;view=1up;seq=357
xlviiWade, A, (April 2020b). Early forays into two-queen hive management: Part I – A plurality of queens. https://actbeekeepers.asn.au/bee-buzz-box-april-2020-early-forays-into-two-queen-hive-management-part-i-a-plurality-of-queens/
xlviiiHall, C.A. (1908). Introducing queens: A modification of the Alexander method. Gleanings in Bee Culture 35(24):1592. https://babel.hathitrust.org/cgi/pt?id=umn.31951d00953180r;view=1up;seq=930
xlix Hand, J.E. (1907). The plural queen system: No problem to introduce a number of queens to bees, but difficult to introduce them to each other. Gleanings in Bee Culture 35(24):1592. https://babel.hathitrust.org/cgi/pt?id=umn.31951d00953180r;view=1up;seq=813
Sherrod, J. (1907). The plural-queen system. More honey than for the one-queen system. Gleanings in Bee Culture 35(21):1593-1594. https://www.biodiversitylibrary.org/item/74679#page/1609/mode/1up
Chambers, J.E. (1907). Two or more laying queens in one hive: The plan not a success in the hands of the average bee-keeper. Gleanings in Bee Culture 35(17):1146-1147. https://babel.hathitrust.org/cgi/pt?id=umn.31951d00953180r;view=1up;seq=670;skin=mobile
Wright, A.J. (1907). The Alexander plan of building up weak colonies, and a modification of it: Two queens in a hive as a method of preventing swarming. Gleanings in Bee Culture 35(21):1886. https://babel.hathitrust.org/cgi/pt?id=umn.31951d00953180r;view=1up;seq=814
Hand, J.E. (1908a). The plural-queen system: Why is it more practicable with a division board-chamber than with an ordinary full-depth hive. Gleanings in Bee Culture 36(1):35-36. https://babel.hathitrust.org/cgi/pt?id=uc1.b3458202;view=1up;seq=49
Hand, J.E. (1908b). The two-queen system: This plan makes it possible to keep the brood chamber packed with brood during the flow: Forcing honey into the supers: Wintering two queens in one hive not desirable. Gleanings in Bee Culture 36(3):155-156. https://babel.hathitrust.org/cgi/pt?id=uc1.b3458202;view=1up;seq=167
Bussy, E. (1908). The plural queen system: A series of interesting experiments: Clipping the queen’s stings so they can’t kill eachother: Do the bees take a hand in royal combat? Gleanings in Bee Culture 36(3):156-157. https://babel.hathitrust.org/cgi/pt?id=uc1.b3458202;view=1up;seq=168
Gray, J. (1908). The plural-queen system: How an English expert looks at the question;: advantages and disadvantages. Gleanings in Bee Culture 36(3):157. https://babel.hathitrust.org/cgi/pt?id=uc1.b3458202;view=1up;seq=169
Hand, J.E. (1908c). The dual and plural queen systems: Conditions under which they may be used: Review of the whole question. Bee World 36(8):507-508. https://babel.hathitrust.org/cgi/pt?id=uc1.b3458202;view=1up;seq=517
Joice, G.W. (1911). Wintering a surplus of queens in one colony: The plan a success. Gleanings in Bee Culture 39(7):221. https://babel.hathitrust.org/cgi/pt?id=uc1.b3458208&view=1up&seq=255
Joice, G.W. (1911). More about wintering a surplus of queens in one colony: Is it worth a trial. Gleanings in Bee Culture 39(14):436-437. https://babel.hathitrust.org/cgi/pt?id=uc1.b3458209&view=1up&seq=100
lMarchant, A.B. (1911). Raising queens above perforated zinc: Plurality of queens for one colony. Gleanings in Bee Culture 39(7):227. https://babel.hathitrust.org/cgi/pt?id=uc1.b3458208&view=1up&seq=261 [Note: Marchant, though an exemplary apiarist, was unable to establish two-queen colonies.]
liWade, A. (May 2020c). Early forays into two-queen hive management: Part II – The two-queen heather honey flow system. https://actbeekeepers.asn.au/bee-buzz-box-may-2020-early-forays-into-two-queen-hive-management-part-ii-the-two-queen-heather-honey-flow-system/
liiRoot, E.R. (1911). Bee-keeping on the Appalachicola River, Florida. Gleanings in Bee Culture 49(24):4751-752. https://ia800905.us.archive.org/8/items/gleaningsinbeecu39medi/gleaningsinbeecu39medi.pdf
Root, E.R. (1912). Bee-keeping on the Appalachicola River. Gleanings in Bee Culture 50(2):46-49. https://ia802702.us.archive.org/24/items/gleaningsinbeecu40medi/gleaningsinbeecu40medi.pdf
Root, E.R. (1912). Bee-keeping on the Appalachicola River. Gleanings in Bee Culture 50(3):80-82. https://ia802702.us.archive.org/24/items/gleaningsinbeecu40medi/gleaningsinbeecu40medi.pdf
liiiSpoja, J. (1953). Observations on the operation of multiqueen colonies. Bee World 34(10):195-200. https://doi.org/10.1080/0005772X.1953.11094822
livKovtun, F.N. (1949). How to make and use multiple queen colonies. Pchelovodstovo 26(9):29-30. Cited by Spoja (1953) and by Latif, A., Qayyum, A. and ul Haq, M. (1960). Multiple and two-queen colonies of Apis indica F.. Bee World 41(8):201-209. Published online: 31 Jul 2015. https://doi.org/10.1080/0005772X.1960.11096796
Kovtun, F.N. (1950). Letter to the editorial office (multiple-queen colonies). Pchelovodstovo 27(2):112. Cited by Spoja (1953). loc.cit.
Also cited by Darchen and Lensky (1963) as Kovtun, F. N. (1949). Comment créer et utiliser des colonies à plusieurs reines? Pchelovodstvo 26:413–41
Kovtun, F. N. (1950). Colonics à plusicurs reines. Pchelopodstvo 27:112.
lvFarrar. (1953). p.189. loc. cit.
lviWade (2019a). loc. cit.
lviiLensky, Y. and Golan, Y. (1966). Honeybee populations and honey production during drought years in a subtropical climate. Scripta Hierosolymitana 18:27-42. in Studies in Agricultural Entomology and Plant Pathology, Avidov, Z. (Ed.). Jerusalem 1966 at the Magnes Press, The Hebrew University. https://catalogue.nla.gov.au/Record/2752040?lookfor=scripta%20hierosolymitana%2
lviiiPaleolog, J. (2001). An attempt at overwintering sting-clipped queens in multiple-queen colonies. Journal of Apicultural Science 45(1):13-20. http://www.jas.org.pl:81/An-attempt-at-overwintering-sting-clipped-queens-in-multiple-queen-colonies,0,7.html
Paleolog, J., Kasperek, K. and Lipiński, Z. (2011). The psychological dimension of duels between western honeybees with blunted and non blunted sting. Journal of Apicultural Science 55(2):85-95. https://www.researchgate.net/publication/286056166_the_psychological_dimension_of_duels_between_westerm_honeybee_queens_with_blunted_and_non_blunted_stings
Siuda, M., Wilde, J., Woyke, J., Jasiński, Z. and Madras-Majewska, B. (2014). Wintering reserve queens in mini-plus and 3-comb nuclei. Journal of Apicultural Research 50(1):61-68. https://content.sciendo.com/downloadpdf/journals/jas/58/1/article-p61.xml
lixZheng, H.Q., Jin, S.H., Hu, F.L., Pirk, C.W.W. and Dietemann, V. (2009a). Maintenance and application of multiple queen colonies in commercial beekeeping. Journal of Apicultural Research 48(4):290-295. https://www.tandfonline.com/doi/abs/10.3896/IBRA.1.48.4.10 and https://www.researchgate.net/publication/229087767_Maintenance_and_application_of_multiple_queen_colonies_in_commercial_beekeeping
Zheng , H.Q., Jin, S.H., Hu, F.L. and Pirk, C.W.W. (2009b). Sustainable multiple queen colonies of honey bees, Apis mellifera ligustica. [Colonias sostenibles de abejas Apis mellifera ligustica con múltiples reinas.] Journal of Apicultural Research 48(4):284-289. https://doi.org/10.3896/IBRA.1.48.4.09 and https://www.tandfonline.com/doi/pdf/10.3896/IBRA.1.48.4.09?needAccess=true
Zheng , H.Q., Jin, S.H., Pirk, C.W.W., Dietemann, V., Crewe, R. and Hu, F.L. (2009c). Honeybee multiple queen colonies in China. https://www.apimondia.com/congresses/2009/Technology-Quality/Symposia/Ten%20years%20of%20beekeeping%20with%20multiple-queen%20colonies%20in%20China%20-%20ZHENG%20Huo-Qing.pdf
Zheng, H.Q., Jin, S.H., Hu, F.L., Pirk, C.W.W., Dietemann, V. (2009d). Maintenance and application of multiple queen colonies in commercial beekeeping. Journal of Apicultural Research 48(4):290–295. https://www.tandfonline.com/doi/abs/10.3896/IBRA.1.48.4.10
Zheng, H.Q., Dietemann, V., Hu, F.L., Crewe, R. M. and Pirk, C.W.W. (2012). A scientific note on the lack of effect of mandible ablation on the synthesis of royal scent by honeybee queens. Apidologie 43(3):471–473. https://hal.archives-ouvertes.fr/hal-01003538/document
Kelsall, A. (1940). A multiqueen colony of bees. American Bee Journal 80:170. Cited by Darchen, and Lensky (1963) and by Riedel and Blum (1972). loc. cit.
Davenes, H. (1907). The plural queen system: The colonies more uniformly strong at the beginning of the honey-flow: Swarming easier to control: Why only one queen is left when excluders are removed. Gleanings in Bee Culture 35(10):1578-1579, 1582. https://ia802703.us.archive.org/8/items/gleaningsinbeecu35medi/gleaningsinbeecu35medi.pdf
lxMostafa, S.N., Magda H.A.S., and El-Ansary, O. (2017). Effect of the multiple queens within colony on some honeybee activities, Apis mellifera carnica and sustainability of their colonies. Mansoura Journal of Plant Protection and Pathology 8(6):277-281. http://main.eulc.edu.eg/eulc_v5/Libraries/UploadFiles/DownLoadFile.aspx?RelatedBibID=NGQ5MGFjMzYtNWE2YS00MGIxLTk3NjctNDNjYzdmNGQ3MWVhX2l0ZW1zXzEyNDkyNTM3XzM1Mjg4OV9f&filename=8-309.pdf
Riedel, S.M. and Blum, M.S. (1972). Rapid adaptation by paired queens of the honey bee, Apis mellifera. Annals of the Entomological Society of America 65(4):825–829. https://doi.org/10.1093/aesa/65.4.825
lxiLensky, Y. and Darchen, R. (1963). Étude préliminaire des facteurs favorisant la création de sociétés polygynes d’Apis mellifica. [Preliminary study of the factors promoting the formation of polygynous societies of Apis mellifica var. ligustica.] Annales de l’Abeille 6(1): 69-73. https://hal.archives-ouvertes.fr/hal-00890173/document
Darchen, R. and Lensky, Y. (1963). Quelques problèmes soulevés par la création de sociétés polygynes d’abeilles. Insectes Sociaux 10(4):337-357. https://link.springer.com/article/10.1007/BF02223064
Lensky, Y; Darchen, R. and Levy, R. (1970). L’agressivité des reines entreeles et des ouvrières vis-a-vis des reines lors de la création des sociétés polygynes d’Apis mellifera L. Revue du Comportement Animal 4(4):50-52. 395/72. Cited by Pflugfelder, J. and Koeniger, N. (2003). Fight between virgin queens (Apis mellifera) is initiated by contact to the dorsal abdominal surface. Apidologie 34:249-256. https://doi.org/10.1051/apido:2003016 and https://hal.archives-ouvertes.fr/hal-00891776/document
Lensky, Y., Cassier, P., Rosa, S. and Grandperrin, D. (1991). Induction of balling in worker honeybees (Apis mellifera L.) by stress pheromone from Koschewnikow glands of queen bees: Behavioural, structural and chemical study. Comparative Biochemistry and Physiology – Part A: Physiology 100(3):585-594. https://doi.org/10.1016/0300-9629(91)90374-L
Mel’Nik, M.I. (1951). Managing multiple queen colonies. Pchelovodstvo 28(9):36-37. Referenced by Zheng et al (2009a). loc. cit.
lxiiChambers, J.E. (1910). Requeening by superseding the old queen in a natural way. Gleanings in Bee Culture 37(6):178. https://babel.hathitrust.org/cgi/pt?id=uc1.b3458204&view=1up&seq=370
Miller, A.C. (1913). Requeening without dequeening: Some interesting facts. Gleanings in Bee Culture 41(23):851-852. https://ia902607.us.archive.org/18/items/gleaningsinbeecu41medi/gleaningsinbeecu41medi.pdf
Miller, A.C. (1914). Why the smoke method of introducing is successful: Requeening without dequeening. Gleanings in Bee Culture 42(14):537-538. https://ia600700.us.archive.org/35/items/gleaningsinbeecu42medi/gleaningsinbeecu42medi.pdf
Hand, J.E. (1914). Requeening without dequeening. Gleanings in Bee Culture 42(1):292.
Smedley, N. (1914). Requeening without dequeening in New Zealand: A duel between two queens. Gleanings in Bee Culture 42(1):292-293.
lxiiiWoyke, J. (1988). Problems with queen banks. American Bee Journal 128(4):276-278. https://www.researchgate.net/publication/256088341_1987_Problems_with_Queen_Banks
lxivSzabo, T.I. (1975). Overwintering of honeybee queens 1. Maintenance of honeybee queens in solitary confinement. Journal of Apicultural Research 14(2): 69-74. https://doi.org/10.1080/00218839.1975.11099805
Szabo, T.I. (1977). Overwintering of honey bee queens: 2. Maintenance of caged queens in queenless colonies. Journal of Apicultural Research 16(1):41-46. https://doi.org/10.1080/00218839.1977.11099858
Wyborn, W.H. (1991). Mass storage of honey bee queens over winter. MSc Thesis, 130 pp. Simon Fraser University. https://summit.sfu.ca/system/files/iritems1/4760/b14477099.pdf
Wyborn, M.H., M.L. Winston, M.L. and P.H. Laflamme, P.H. (1993). Mass storage of honey bee (Hymenoptera: Apidae) queens during the winter. The Canadian Entomologist (125):113-128. https://doi.org/10.4039/Ent125113-1 Gençer, H.V. (2003). Overwintering of honey bee queens en mass in reservoir colonies in a temperate climate and its effect on queen performance. Journal of Apicultural Research 42(40):61-64. https://www.researchgate.net/publication/247161387_Overwintering_of_honey_bee_queens_en_mass_in_reservoir_colonies_in_a_temperate_climate_and_its_effect_on_queen_performance
http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.632.753&rep=rep1&type=pdf
Shehata, S.M. (1982). Long-term storage of queen honeybees in isolation. Journal of Apicultural Research 21(1):11-18. https://www.tandfonline.com/doi/abs/10.1080/00218839.1982.11100510
Reid, M. (1975). Storage of queen honeybees. Bee World 56(1):21-31. https://doi.org/10.1080/0005772X.1975.11097534
Klassen, S., Guzman, E. and Kelly, P. (2017). Banking multiple queens in colonies overwinter. Ontario Bee Journal January/February:12-13. Accessible as Klassen, S., Guzman, E. and Kelly, P. (2015-2016). Experimental queen overwintering study at University of Guelph (2015-2016) in Bixby, M., Guarna, M., Hoover, S.E. and Pernal, S.F. (2018). Canadian honey bee queen bee breeders’ reference guide, pp.39-40. http://honeycouncil.ca/wp-content/uploads/2018/12/FinalQueenBreederReferenceGuide2018.pdf
Harp, E.R. (1967). Storage of queen bees. American Bee Journal 107(7):250-251.
Harp, E.R. (1969). A method of holding large numbers of honey bee queens in laying conditions. American Bee Journal 109(9):340-341.
Levinsohn. M. and Lensky, Y. (1981). Long-term storage of queen honeybees in reservoir colonies. Journal of Apicultural Research. 20(4):226-233. https://doi.org/10.1080/00218839.1981.11100501
lxvDietz, A., Wilbanks, T.W. and Wilbanks, W.G. (1983). Investigations on long-term queen storage in confined systems. Apiacta 3:1-3. http://www.fiitea.org/foundation/files/1983/A.%20DIETZ,%20T.W.%20WILBANKS,%20W.G.%20WILBANKS.pdf
Dietz, A. (1985). Problems and prospects of maintaining a two-queen colony system in honey bees throughout the year. American Bee Journal 125(6):451-453.
lxviGary, N.E. (1963). Observations of mating behaviour in the honeybee. Journal of Apicultural Research 2(1):3-13. https://doi.org/10.1080/00218839.1963.11100050
Gary, N.E. (1966). Maintenance of isolated queen bees under laboratory conditions: A preliminary research report. American Bee Journal 106(11):412-414. Cited by Reid (1975). loc. cit.
Gary, N.E., Hagedorn, H.H. and Marston, J. (1967). The behavior of mated queens when colonized in multiple queen groups without worker bees. Apiacta 3(4):9-12. http://www.fiitea.org/cgi-bin/index.cgi?sid=&zone=cms&action=search&categ_id=51&search_ordine=descriere Gary, N.E. (1959). A study of natural and induced supersedure of queen honeybees (Apis mellifera L.). PhD thesis, Cornell University, Ithaca, N.Y., A.A.586/64, 145pp. https://www.researchgate.net/publication/34234849_A_study_of_natural_and_induced_supersedure_of_the_queen_honey_bee_Apis_mellifera_L
lxviiSzabo, T.I. and Townsend, G.F. (1974). Behavioural studies on queen introduction in the honeybee 1: Effect of the age of workers from a colony with a laying queen on their behaviour towards an introduced virgin queen. Journal of Apicultural Research 13(1):19-25. https://doi.org/10.1080/00218839.1974.11099754
Szabo, T.I. (1974). Behavioural studies on queen introduction in the honeybee 2. Effect of age and storage conditions of virgin queens on their attractiveness to workers. Journal of Apicultural Research 13(2):127-135. https://doi.org/10.1080/00218839.1974.11099768
Szabo, T.I. (1974a). Behavioural studies on queen introduction in the honeybee 3: Relationship between queen attractiveness to workers and worker aggressiveness towards a queen. Journal of Apicultural Research 13(3):161-171. https://doi.org/10.1080/00218839.1974.11099774
Szabo, T.I. (1974b). Behavioural studies on queen introduction in the honey bee. 4: Introduction of queen models into honey bee colonies. American Bee Journal 114(5):174-176.
Szabo, T.I. and Smith, M.V. (1972). Behavioural studies on queen introduction in honeybees. 5: Behavioural relationship between pairs of queens without worker attendance. Proceedings of the Royal Society of Ontario 103:87-96. Findings cited by Pflugfelder, J. and Koeniger, N. (2003). Fight between virgin queens (Apis mellifera) is initiated by contact to the dorsal abdominal surface. Apidologie 34(3):249-256. https://hal.archives-ouvertes.fr/hal-00891776/document
Szabo, T.I. (1977). Behavioural studies of queen introduction in the honeybee 6: Multiple queen introduction. Journal of Apicultural Research 16(2):65-83. https://doi.org/10.1080/00218839.1977.11099865
Waikakul, Y. (April 1979). Behavioral studies of virgin queen honey bees. MSc Thesis, The University of Manitoba, Winnipeg, Manitoba. https://www.google.com/search?q=Waikakul%2C+Y.+(May+1973).+Behavioral+studies+of+virgin+queen+honey+bees&oq=Waikakul%2C+Y.+(May+1973).+Behavioral+studies+of+virgin+queen+honey+bees&aqs=chrome..69i57.2013j0j7&sourceid=chrome&ie=UTF-8
lxviiiFarrar (1953). loc. cit.